[拼音]:shenghuohuan(dongwu)
[外文]:life cycle (animal)
又称生活周期,即由成年亲体产生的受精卵经过胚胎发育作为新一代个体诞生,自幼而长,达到性成熟时又具备了生殖能力的全过程。有机体生灭相继,形成无尽的生命环。其自我复制能力是维持生命环延绵的基础,而生活环则体现为复制的具体过程。
生活环大致包括生殖、胚胎发育和生长等阶段。实际上这并非生物个体的全部生活历程,因为未把衰老包括在内(图1)。20世纪中叶以前常用的生活史一词就有包括衰老在内或不将之包括在内的混统含意。
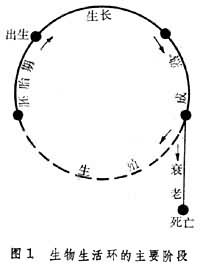
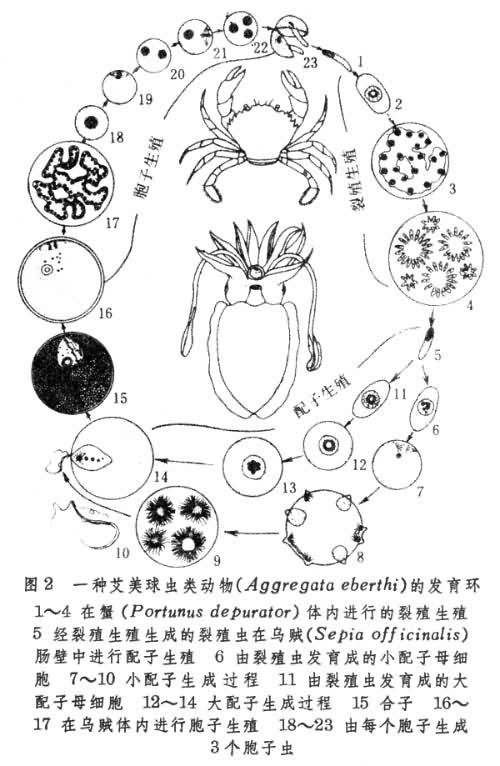
有些动物,特别是低等无脊椎动物的生活环相当复杂。不同生殖方法,不同形态发生和不同生活方式可出现于同一生活环的不同时期,因而使整个环的组成包括几个不同世代。如胞子虫的生活环中就有胞子虫、裂殖虫和配子生成3个世代(图2)。前两者属无性世代,末者属有性世代。由此三者形成的交替或循环又常称为发育环。在这里,发育环所代表的内容比一般生活环中只包括几个发育阶段的情形要复杂些。
性发育环与生活环
动物主要通过有性生殖代代相传。故常以前一代性细胞结合和下一代性腺成熟分别作为生活环的起止点。也可说由性结合到性成熟构成以性腺发育周期为中心内容的性发育环,生活环与性发育环同步进行。
有些动物终生只有一个性发育环,但更多种类具有多个性发育环。在前种情形下性发育环几乎等同于生活环;在后种情形下,重复的性发育环当然要超出了生活环的原定范围。此外,较特殊的是有些兜水母类动物(Eu-charis multicornis,Bolina hydatira)一生中限有两次性发育环:一次在幼虫期,另一次在成体期。其间幼虫必须排完精卵之后才能变态为成体,在成体期间再完成一次性发育环。这种具有两次性发育环的生活环又称重复生殖。
与性发育环相联系的另一个发育生物学概念是生殖环或生殖周期。由于有一个性发育环就会有一个生殖环,所以后者既是前者的目的,也是前者的结果。根据历经生殖环次数的不同(实际上也就是性发育环次数的不同),动物生活环可以分为两种:单殖生活环和复殖生活环(图3)。
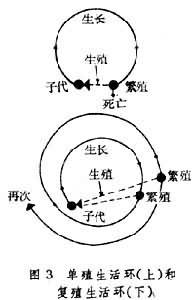
动物个体长成后只进行一次有性生殖即行死亡(某些昆虫)的属前者,个体长成后重复生殖的属后者。这是动物界生殖上的两种策略。用一次多产的办法或用多次少产的办法都有可取之处,也都具弱点。新生子代能活到性成熟期的几率和亲体历经各生殖时期的存活率都是复殖生活环成效的重要依据。由于成体对环境的适应能力总比幼体为强,所以单殖生活环不如复殖生活环优越。
关于性发育对生活环内容形式所起的作用,从水螅虫类中可以找到较恰当的例子。如有种水螅虫(Coryden-drium parasiticum)的原始生殖细胞在发生过程中由中胶层迁入外胚层,结果使所移居位置变成了生殖芽体的生成位置。这表明芽体的生成是受生殖细胞的诱导。另一个例子是从众多生殖芽体类型的比较中看到一个现象:生殖细胞的发生越早,其所在生殖芽体的形态分化程度越差。如在茎突型生殖芽体(又称胞子囊)发生过程中,所有伞体器官都不出现,而生殖细胞却在一开始就分化了出来(图4)。
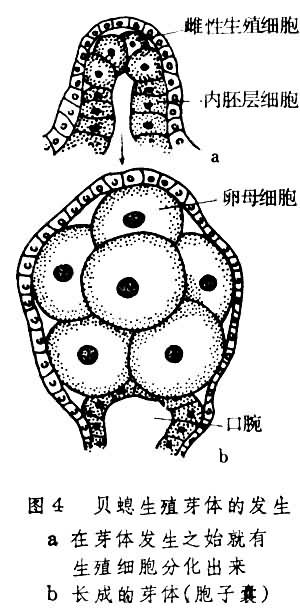
在这种情形下,该型芽体竟与淡水水螅生殖腺相差不远,事实上已不足作为有性世代个体。以上两例共同说明:不论是生殖芽体的生成或是此芽体的退化都与当时性发育情况有联系。水螅虫生活环中有无水母体世代出现可能就是这种联系的结果。
有世代交替的生活环
主要的和常见的两种是:无性世代与有性世代相交替的生活环和单性世代与两性世代相交替的生活环。后者又称易性交替生活环。由于单性世代和两性世代各对环境有一定要求,所以易性交替生活环具有较复杂的过程。如有种蚜虫(Sacchiphantesviridis)先以云杉为食,后改为以落叶松为食。由其性蚜期产生的受精卵在冬日孵化为建群个体。春夏间由建群个体产生的新一代为翅蚜,或称迁移性蚜。这种翅蚜在找到落叶松后再产出处女蚜。以落叶松为食的处女蚜可历经多个世代,直到过冬后才又生出另一代有翅的产性蚜。这种蚜虫分为生雄蚜和生雌蚜。它们都移食于云杉并各继续孤雌生殖直至分别生出雌、雄性蚜。经过交配和产卵之后性蚜死亡,由受精卵再孵出建群个体。是为新一代生活环的开始。上述蚜虫生活环中包括多个生殖环的意义主要在利用不同世代来渡过不同环境,以保种的生存。这和甲壳类动物生活环中利用多个幼虫期来渡过不同时期的环境有异曲同工之妙。
有变态的生活环亦称变态型生活环。多种无脊椎动物和很少数脊椎动物有此型生活环。这种生活环的特点在于具有双重发育体制:先是幼虫期,然后是成体期。由前者向后者的过渡要经历一种加速程序。其中既有某些幼虫器官的退化或破坏,同时又有新组织和成体器官的生成。纽虫、苔藓虫和昆虫的坏死性变态是其中最突出的例子。
变态现象的产生与祖体动物生活方式的剧烈变化有着密切的关系。如由水生生活改为陆生生活(蛙),由浮游生活改为底栖生活(海胆)以及由步行生活改为飞行生活(昆虫)等势必要求有关动物的某些结构及其技能作相应而迅速的变化。变态就是这种历史性重大事件在生活环中的反映。
在变态型生活环中,受精卵、幼虫期和成体期的基因组是相同的。幼虫和成体间所以有形态差别则是由于它们一先一后受不同基因型的支配而显示出的两个不同的表型。可把幼虫形态看成是原始表现型,成体形态是后生表现型。
季节环或季节性周期有很多昆虫从某季节到翌年同一季节形成一个生活环,但也有不少种类可形成多个生活环,这两种情形统被称为一个季节环。
在昆虫中,生活环历时超过一年的情形也是存在的。如六月鳃金龟子的幼虫期普遍为2~3年,多种蝉类动物的幼虫期为2~5年,有的Magicicada长达13或17年。
在季节环概念下,有的昆虫虽具多个生活环,但情况仍较简单。如经过冬眠后的家蝇入夏后可连续出生多个世代,每隔4~5周就完成一个生活环并且各环过程相同。但是更有不少种类的季节环却相当复杂。例如甘兰蚜在秋季产出的越冬卵于次年春孵育为无翅、能进行孤雌胎生的个体,即是其建群个体。之后,由此种个体所递传下来的夏季各代个体依然进行孤雌胎生;必须到秋季始能胎生出有翅雄体和无翅雌体。经交配后,每个雌体能产 1至数个受精过的越冬卵。以上表明甘兰蚜的季节环和前述蚜虫的一样,既包括了多个世代,同时也因单性世代和两性世代先后出现而构成世代交替。不同之处在于:甘兰蚜在季节环中并无改换寄主的情形,而另种蚜虫却有迁徙世代的出现而达改换寄主的目的。
在昆虫中还有一种形式很特殊的季节环,即:出现在此环中的许多世代并不是成体而是幼虫,由幼虫产生幼虫。至于何时由幼虫生成两性成体询以客观环境条件而定。这种情形可称之为幼体生殖式的季节环。
环境条件对生活环的影响
光线对某些节肢动物的滞育和蜕皮有特别明显作用,温度则对动物的休眠(冬眠、夏眠)、迁徙,洄游和生殖有很大影响。有种笔螅(Pennaria tionella) 的水母体世代可随季节水温变化而有形态和生态上的变化。当春季水温尚低时,其水母体既有较完整的形态又能自由生活;但夏季水温升高后,不仅形态退化,并且也不再离开水螅体。
水域和底质的环境常是决定底栖动物生存和分布的关键条件。有种前鳃动物(Colus islandicus)在某海区中发育时有浮游幼虫期出现,而在另一海区中却无此幼虫期。有种海星(Mediaster aequalis)的幼虫在找不到一种多毛类动物栖管时,可长时期不沉落,也不进行变态,必需在获得必要条件后才能完成其生活环中的后期阶段。
各种寄生动物都有特殊的寄生方式借以应付各自特殊环境,从而导致其生活环的多样化。如线形动物的生活环就有近20种类型。它们的区别在于有的种类完全营寄生生活,有的营半寄生生活;有的在发育早期(又分不同幼虫期)进入寄主,有的在成体期才进入寄主;有的寄生历时较长,包括幼虫期和成体期,有的历时较短,限某一幼虫期或成体期;有的需要单一寄主,有的在不同发育期需要不同寄主,如终局寄主、中间寄主或传送寄主等。
寄生动物经长期演化过程而使其生活环各环节高度地适应寄主生活环中的各环节。如有种寄生在蠊或白蚁中的鞭毛虫(Crytocercus punctulatus),其性发育周期与寄主蜕皮周期同步进行。如果寄主因受处理而有减慢或停止蜕皮的情况,其性发育环亦即作相应改变。在这里究竟是蜕皮本身产生的影响还是寄主激素产生的影响尚不清楚。
除上述外,某些化学物质及其浓度的变化也可成为使动物生活环中某阶段产生敏感性影响的环境条件。如氯化汞浓度升高可引起海胆的先期变态,碘、铁、铝等离子在无毒浓度下能引起海鞘幼虫的变态,而冬春季节海水中碘浓度的提高则能引起海月水母进行横裂生殖。
某些化学物质对原生动物的性发育所产生的影响也很明显。如水中缺氮,衣藻即停止无性分裂而产生配子。另一种鞭毛虫 (Duneliella salina)在盐度降低时也会出现上述情形。
生活环的逆转现象
此现象见于水母类、纽形动物、海鞘类和扁形动物三肠类等许多物种。经过长期饥饿后,它们缩小身体或作倒退生长以致无论在形态上或是在生理上都有还童表现。根据辐射实验,有人认为三肠类动物是因有种致老因子不断在体内积累以致变老。而在倒退生长中很可能该因子发生异化分解,以后在重新生长中由于其密度的减低,所以发生还童现象。有的学者提出:在这些动物还童过程中可能有成体未分化细胞在起作用。这种细胞经常处于潜隐状态,但在动物个体受过饥饿而又重获营养时即进分裂,产出有较强代谢能力的幼细胞。由后者再分化为他种体细胞。
形态演化在生活环中的体现
在各类动物生活环中所出现的形态发生和变化,不仅反映了物种的历史和演化水平,同时也是它们各自演化途径的记载。在漫长而艰难的岁月中,它们的形体在不断地适应环境中得到改进或发展。这种改进或发展建立在原有基础上,既可实现于某一系统或器官,又可实现于生活环的任一阶段。下面的几种形态演化方式可以作为说明。
幼态持续现存动物在成体期出现祖体动物幼虫期形态的现象。也有人认为是幼虫期即达到性成熟的现象。如海鞘中的幼形目动物,一方面保留着幼虫尾,一方面又是性成熟个体。
新性发生为应付环境需要,在胚胎或幼虫期产生暂时性器官的情形。如担轮幼虫的纤毛轮和摇尾幼虫的尾作为运动器官都属这类构造,后在变态中退化消失。
幼体形态发育出于系统发育中保留下来的幼虫期提前性成熟,于是后期形态发生便不再按原有程序进行,即演化方向有了变更。如有种寒武纪棘皮动物的浮游性幼虫与现存棘皮动物者极为相似,只是前者因为提前性成熟便不再沉落,也不再发育为营底栖生活的成体,转而开始了新的形态发育。
退化祖体动物成体或幼体有过的构造消失于现存动物的成体期的现象。有些动物(扁形、线形)是因为营寄生生活有这种情形,另有些腔肠动物受饥饿后也会有这种情形。
附加发育有些器官或构造在祖体动物时期未曾出现或得到发展,而是在后代动物中才出现或得到发展。如藤壶幼虫是该动物在系统发育中获得的形态,同时也是多类甲壳动物共有的形态。但是其附着后所出现的形态却是他类相近动物所没有。从意义上来说,这就是该动物在演化过程中获得的附加形态,当然也就成了生活环中的新内容。
又如雷兽的角和犬齿虎的犬齿都在演化中有过分的发展,从而也具附加发育意义。
延迟发育有些现存动物的器官与祖体动物者相比较,从一开始就推迟了发生时间或是减低了发育速度。如人类的智齿。
研究意义
生活环研究与生产密切相关。最早的研究对象是桑蚕。中国荀卿在纪元前三世纪就在《蚕赋》中记载了该动物“三眠三起,事乃大己”的生活过程。在欧洲,17世纪荷兰人J.斯瓦默达姆明确了蝇、蝶的变态型生活环。但广泛地科学地开展生活环研究则是从19世纪中叶才开始,在20世纪内有了很大进展。其成果已应用于许多方面,如:人、畜寄生虫病的防治,农业害虫的消灭,有益动物和食用动物的养殖和保护,附着动物的防除,环境污染的监测以及生物防治中对食害动物的利用等。今后为进行动物区系改造和野生动物保护等工作也有大力开展生活环方面研究的必要,因为这是了解某一地区范围内食物链情况所不可缺少的基础。