[拼音]:caoyuan shengtai
[外文]:steppe ecology
草原上的生物之间及生物与环境之间的关系。草原是内陆半干旱到半湿润气候条件下所特有的一种自然生态系统类型。一般辽阔无林,旱生多年生禾草占绝对优势,多年生杂草及半灌木也起显著作用;通常栖居着各种善于奔驰或营穴洞生活的哺乳动物,以及其他动物。
草原概述
世界草原总面积约2400万平方公里,为陆地总面积的1/6,大部分用作天然放牧场。
草原可分为温带草原与热带草原两类。温带草原分布在南北两半球的中纬度地带,如欧亚大陆草原、北美大陆草原、南美草原等。这里夏季温和,冬季寒冷,春季或晚夏有一明显的干旱期。由于低温和雨量较少,草群较低,其地上部分高度多不超过 1米,以耐寒的旱生禾草为主。土壤中以钙化过程和生草化过程占优势。分布在热带、亚热带的草原,其特点是在高大禾草(常达2~3米)的背景上常散生一些不高的乔木,故被称为稀树草原。这里终年温暖,雨量常达1000毫米以上。在高温多雨影响下,土壤强烈淋溶,以砖红壤化过程占优势,比较贫瘠。一年中存在一个到两个干旱期,加上频繁的野火,限制了森林的发育。
草原是一种地带性的生态系统类型。地球上的草原虽然从温带分布到热带,但在气候坐标轴上却占据固定的位置。在寒温带,年雨量150~200毫米地区已有大面积草原分布,而在热带,稀树草原却主要分布在 800~1400毫米雨量的地区,前者的低温少雨与后者的高温多雨产生相似的生物学效果。因此,水分与热量的组合状况是影响草原分布的决定因素。在地理分布上,草原处于湿润的森林区与干旱的荒漠区之间。靠近森林一侧,气候半湿润,草木繁茂,种类丰富,并常出现岛状森林或灌丛,如北美的高草草原、欧亚大陆的草甸草原以及非洲的高稀树草原。靠近荒漠一侧,雨量减少,气候变干,草群低矮稀疏,种类组成简单,并常混生一些旱生小半灌木或肉质植物,如北美的矮草草原、中国及蒙古的荒漠草原以及苏联欧洲部分的荒漠草原等。在上述二者之间为辽阔而典型的禾草草原(图1)。
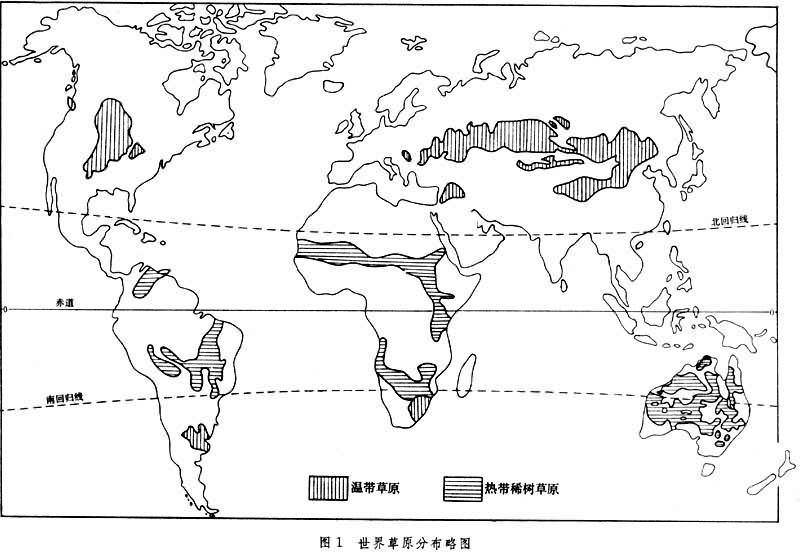
总的看来,草原因受水分条件的限制,其动植物区系的丰富程度及生物量比森林低,比荒漠高,但与森林和荒漠比较起来,草原动植物种的个体数目以及较小单位面积内种的饱和度是相对丰富的。
草原动植物区系
作为生产者的绿色植物是草原生态系统组成的基础,草原植物或多或少具旱生特征。其基本生活型为地面芽植物,尤其是丛生禾草,是草原群落的主要建造者。地下芽植物在草原上也起着重要作用,其中如根茎禾草与根茎苔草,在不少草原地区(尤其是轻质土壤上)起着优势作用。主要由双子叶植物组成的各种杂类草,在草原上也非常丰富。如菊科、豆科和蔷薇科的一些种,在靠近森林地区即水分条件较好地段繁茂发育,与禾草共同起优势作用,其根系深,能贮藏水分和养分,故也能分布于干旱地区。有些杂类草的根系伸至禾草根系层之下才开始分枝,其深度可达禾草根系深度的2倍(图2)。

至于地上芽植物,主要是不高的灌木与半灌木,在草原上多少起显著作用,尤其在接近荒漠地区,它们的作用大大增强,有时可达优势地位,如欧亚大陆草原的锦鸡儿、亚菊、蒿、地肤等属的一些种。高位芽植物中的乔木,在温带草原中极为少见,但广泛分布于热带稀树草原中,如非洲的阿当松氏金合欢及大洋洲的桉树等,均具顶部平展的伞状树冠。种子越冬的一年生植物在草原上的作用不大,但在具地中海气候型的草原地区,有一类短生植物,常在春季繁茂发育,如球茎早熟禾、二花郁金香等。苔藓、地衣和藻类也是草原上常见的植物类群,它们居于地表,只能利用土面和土壤表层的水分和养分。
生态学家常按高度把草原上占优势的多年生草本植物分为 1米以上的高草、30~90厘米的中草和30厘米以下的矮草。高草主要分布在水分条件较好的热带、亚热带稀树草原以及温带半湿润地区,多为中旱生植物甚至中生植物;矮草分布在靠近荒漠一侧的干旱地区,形成矮草草原和荒漠草原,多为典型的旱生植物和强旱生植物;中草的分布介于二者之间,为典型草原的主要成分。
根据草原植物对温度的反应,可分出耐寒型和喜暖型两大类。耐寒型多分布在中温带、寒温带及高寒山地草原群落中,以早熟禾系的禾草为主,如针茅、羊茅、早熟禾、冰草、隐子草等属的一些种,它们多为呌植物;喜暖型则分布在热带、亚热带稀树草原以及温带草原的暖温带地区,以黍系禾草为主,如黍、狼尾草、芒须草、扭黄茅、萑稗、高粱等属的一些种,它们多为C4植物。
不同地区草原植物种类的丰富程度不同。生态条件越适宜,种类就越丰富,群落结构也越复杂,有地上及地下亚层的分化。反之,生态条件越严酷,种类就越简单,群落结构也越简化。如中国大兴安岭两侧的草甸草原群落,每平方米约有种子植物20~30种或更多;广大的典型草原,每平方米约15~20余种;而干旱的荒漠草原,每平方米仅12种左右。在种的饱和度低、群落结构趋于简化的情况下,地上部分常常不能郁闭,覆盖度多在30%以下,但其地下部分却是郁闭的。草原植物地上生物量与地下生物量的比值随生境的变劣而降低。
在草原群落中起建群作用的植物只是很少数,绝大部分植物为伴生种。如中国草原地区共有种子植物4000种左右,但建群植物只有45~50种,其中主要是禾草,约占全部建群种的75%。由于建群植物的形成历史与生态性质不同,其分布有明显的地域分异,从而形成不同的草原类型。
草原动物区系也很复杂,最引人注目的是大型植食动物,如非洲的斑马、瞪羚、长颈鹿,美洲的野牛,大洋洲的有袋类、麋鹿,欧亚大陆草原的野驴、黄羊等以及牛羊等家畜。此外还有数量众多的其他类群,它们大部分为植食性。植物性食料虽易获得,但营养价值不及动物性食料,必须摄取更大的量才能满足营养要求,因而植食动物在形态上和行为上有一系列的适应。骆驼、袋鼠、长颈鹿、鹿及牛、羊等各自独立地发展了反芻习性。这一特性可以使它们的采食时间缩短,到达安全地区后再将所进食物反芻咀嚼,从而减少了被捕食的机会。营地上生活的草原动物常以快速奔驰保证其安全,常可见到一些善跳善跑类群。善跳的如袋鼠、跳鼠、兔与跳兔,善跑的如羚羊、黄羊、鹿等。它们体轻腿长,异常敏捷,在空旷的草原上能将捕食者远远地抛在后面。鸟类中的鸵鸟、鸸鹋等也属于善跑的一类。有些小型哺乳动物(如啮齿类)则营地下穴洞生活,以适应草原气候的季节变化和逃避捕食者。它们挖掘洞穴,使土壤混合与疏松,在草原群落的形成与演替中起着很大作用。至于大型哺乳动物如象与野牛等,则以其强大的体型为自卫手段。草原动物的视力多敏锐,一些小型哺乳动物在观察周围情况时,常会遇到草丛的障碍,于是其中的一些种类用后腿坐起来窥伺周围环境。多种多样的草原肉食动物,从大型的狮、豹、猎豹到狼、狐、獾及鹰等,它们可以抑制某些草食动物种群数量的大发生,从而维持草原生态系统的稳定与平衡。无脊椎土壤动物的数量更多,它们在草原生态系统中也起着显著的作用。
世界各地草原概述
欧亚大陆草原
地球上最辽阔的温带草原区域,自欧洲多瑙河下游起,呈带状往东延伸,经匈牙利、罗马尼亚、苏联、蒙古人民共和国,直至中国东北平原,然后转向西南,经内蒙古高原、黄土高原达青藏高原的南缘,绵延 8000余公里(图3)。这一区域环绕在亚洲荒漠区的外围,东西两侧与温带落叶阔叶林相接,北连寒温带针叶林。
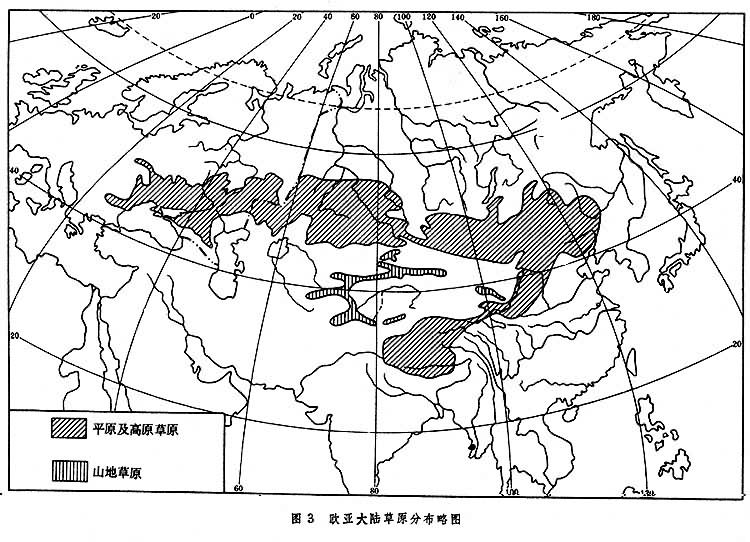
在欧亚大陆草原范围内,水热条件在一年中的分配状况各地有较大差异。西部为地中海型气候,冬春温和多雨,夏季炎热干旱。草原生物于春季繁茂生长,夏季往往进入休眠状态,草群中发育了大量春季短命植物和类短命植物。草原区东部为太平洋季风气候,冬季寒冷少雪,夏季炎热多雨,水热同季,植物生长呈单峰型,夏季最为繁茂,缺少春季短命植物及类短命植物。青藏高原降水分配也为季风型,但气温终年较低,尤其是生长季气温低,因此这里发育了适应于高寒气候的动植物类型,草原中出现垫状植物层片。
除了上述区域差别之外,因水热组合状况不同而引起的地带分异也很明显。在欧亚大陆草原区内,从靠近森林一侧向荒漠区推近时,年降水量逐渐降低,热量渐趋增加,从而引起草原的一系列变化。主要是有效水分越来越少,土壤淋溶程度逐渐减弱,钙积层渐趋于地表,植物高度、密度与种的数目渐趋降低等,动植物区系组成也发生相应的更替,出现森林草原、典型草原与荒漠草原3个自然带的分化。
森林草原带动植物区系丰富,草群密茂,并有岛状桦、杨林及栎林出现。草群下发育了肥厚的黑土、黑钙土及黑垆土。其建群植物西部为约翰针茅、狭叶针茅,蒙古高原东部和中国东北部为贝加尔针茅、中国滨麦及线叶菊,黄土高原为白羊草,草群中混生相当丰富的中生性杂类草。大型草食动物主要有驼鹿、鹿类、麅、原羚等。由于这一地带气候适宜,土壤肥沃,已大部开垦为农田,为春小麦的主要产区。
典型草原带比森林草原带更偏向内陆,具典型的半干旱大陆性气候,发育了栗钙土。草原植物组成简单,以旱生丛生禾草尤其是针茅属占绝对优势,当针茅开花季节(6~8月),其银色的长芒随风波浪起伏,一望无际,十分壮观。西部以几种羽状芒针茅占优势,如列兴针茅、针茅、紫针茅等;东部建群植物以光芒组的几种针茅为主,如大针茅、克氏针茅、长芒草以及糙隐子草、冷蒿等,草群中缺少中生性杂类草。典型草原上常见的草食动物有黄羊、高鼻羚羊以及黄鼠、鼠兔、旱獭等。目前,典型草原地带除局部开垦为农田外,大半保持自然状态,为放牧畜牧业的主要基地。
荒漠草原带从典型草原到荒漠的过渡地带。年降水量仅120~180毫米,而蒸发量却数倍于此值,是草原区内最干旱的地区。草群低矮稀疏,草高仅20厘米左右,覆盖度很低,这为一年生植物的发育提供了空间。每逢多雨年份的雨季,生长起繁盛的一年生植物层片,为草原增添了不少色彩。荒漠草原的群落组成极为特化,如须芒组的小针茅、亚菊等属的小半灌木以及不少啮齿类动物均为这一地带所特有。主要建群植物有新疆针芒、戈壁针茅、短花针茅、蓍状亚菊、女蒿、碱韭以及青藏高原的青藏苔草等。荒漠草原的主要草食动物为沙鼠、短耳仓鼠、高鼻羚羊等。由于蒸发强烈,土壤中的钙及易溶性盐类积累于地表,腐殖质含量低,为棕钙土。这里无灌溉即无农业,但可用作良好的放牧场。
北美大陆草原
分布也极为辽阔,北从加拿大南部起,经美国中部达墨西哥湾;西起落基山东麓,东至美国五大湖区西岸,南北延伸约3700公里,东西宽约700~800公里(图4)。北邻针叶林带,南部逐渐过渡到稀树草原,东接落叶阔叶林,西为落基山山地针叶林及蒿类草地。
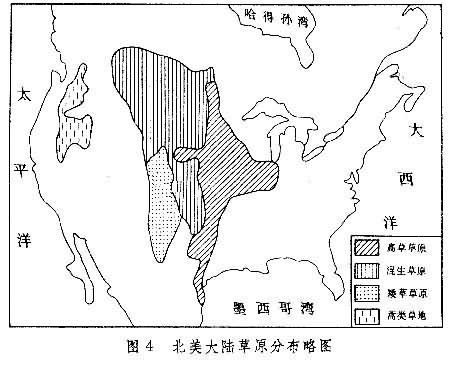
北美草原的分布格局与欧洲大陆有所不同。这里雨量从东往西减少,东部边缘年降水量达 750(北段)~1000毫米(南段),中部500毫米上下,西部降至400毫米以下。与此相应,优势植物的高度从东往西逐渐降低,种类组成也发生明显变化,出现3个地带的依次交替:高草草原、混生草原与矮草草原。从北而南气温逐渐增高,最冷月(1月)平均气温北部低于-20℃,而南部高于0℃,反映在草原组成上也发生明显变化,开花迟的喜暖种在南方占优势,而在北方则生长耐寒种类。
高草草原发育在肥沃的黑土上,原始状态下以须芒草属植物占优势。已多被开垦,种植禾谷类作物及多年生牧草。
混生草原特点是高草与矮草混生,显然是一过渡带。
矮草草原从混生草原带往西南,高草逐渐消失,过渡到矮草草原。这里以野牛草及格兰马草等矮禾草占绝对优势,草群高度30厘米左右,根深达 1米。种的丰富度明显降低,1平方公里仅20种上下,杂类草的数量下降,但混生一些矮灌木、半灌木及肉质植物,如蒿及黄茋等属的一些种以及肉质化的多刺仙人掌。
动物区系北美草原曾经栖息过丰富的动物区系,如古象、乳齿象、古骆驼、野牛、驼鹿等植食动物和以它们为食的美洲豹、美洲狮等猫科肉食动物,但大多已经灭绝,只有野牛与驼鹿由于印第安人家养而存活下来。目前,牛、马、羊等家畜取代了原来的野生动物。
南美草原
主要分布在阿根廷中东部平原,约处于南纬32°~38°之间,面积约50万平方公里。草原区北部降水量达1000~1250毫米,是所有温带草原中降水最高的,往西南雨量渐少,至草原的西南边缘,降水量为500毫米左右。这里气温较高,蒸发强烈,气候相当干燥。当降水量进一步降低,或温度进一步升高时,草原便为旱生灌丛或稀树草原所代替。北部轻质土壤上出现具刺朴树组成的岛状林,优势草原群落主要由奈西针茅和白羊草等禾草组成,混生相当多的杂类草;目前原始群落残存无几,大部已被农田代替。西部及西南部生长了以南美针茅和三歧针茅为主的大型丛生禾草草原,几乎很少杂类草。南美草原上的畜牧业仅有400年的历史,但近一个世纪以来的过度开垦和重牧使生产力持续降低,植被组成也多为引入成分所代替。
稀树草原
主要分布于非洲、大洋洲及南美洲的热带、亚热带半干旱地区。在大洋洲叫scrub,巴西叫cam-pe,南美北部叫llano。后来,南美北部的llano被西班牙人叫做savanna(稀树草原),此后就把类似的草地类型都称之为稀树草原。
非洲的稀树草原分布于赤道雨林的南北两侧,介于北纬15°和南纬25°之间,形成东西延伸的带,北侧带宽400~550公里,东西长约5000公里;南侧带宽200公里,东西长约2500公里。总面积80多万平方公里。优势植物为须芒草族及黍族的高大禾草。散生树木以具刺的金合欢属为主,也有合欢亚科的其他属以及另外一些科的耐旱树木生长,多具刺。在靠近热带雨林一侧分布了以狼尾草属和白茅属为主的高稀树草原,往北或往南,随干旱程度的增加,草群变低矮,以菅草最为丰富。
稀树草原植物的生长期决定于干旱期的长度,一般为120~190天。土壤高度淋溶,以砖红土壤化过程占优势。分解过程快,腐殖质积累量低。经常的火灾对保持稀树草原的稳定起着一定作用;但同时也消灭了对火敏感的种类,促进耐火种类的发展,从而成为稀树草原演替的动力。
非洲稀树草原的动物区系丰富,其中很多为非洲所特有,如非洲象、长颈鹿、斑马、非洲野牛、斑鬣狗、非洲野狗、獴等。此外还有很多小型哺乳动物如啮齿类,数量众多的鸟类(如著名的鸵鸟、鹰等);以及大量的昆虫和其他无脊椎动物。这些动物具有相当高的生物量,如对东非 1.5平方公里稀树草原的估计,有蹄类的生物量每公顷达50千克,小哺乳动物4~5千克,肉食动物8.5千克,无脊椎动物的生物量每公顷可达100千克。
南美稀树草原以委内瑞拉的 llano面积最大,位于安第斯山脉奥里诺科河之间,优势植物为黍属禾草,而具刺树木为高大的仙人掌所取代。这里多被开垦,已很难找到原生植被。
大洋洲稀树草原也有相当大面积,这里仍存在金合欢但却没有具刺的树和灌丛,散生树木的优势树种是桉树,优势草本植物为高大禾草Astrebla属等,雨季繁茂生长,旱季干枯,营养价值很低。特有动物有袋鼠、树熊、袋狼等,其中袋狼可能已近于灭绝。
草原的发生与演变
草原的发生至少可追溯到距今 700万年之前的中新世。从中新世起到第四纪中期,中国地壳曾发生一系列巨大变动。喜马拉雅山、昆仑山、天山、阿尔泰山和青藏高原先后在西部隆起,同时东部临海部分升起许多山地,蒙古高原和黄土高原也在这时形成。地形的升起阻挡了北大西洋和印度洋湿气流东上,同时也减弱了太平洋湿气团的西进,使中国北部日趋寒冷、干旱,促成干旱、半干旱区的形成。中新世时期,中国干旱区大致为稀树草原景观,仅在山地开始形成草原群落片段。到中新世末,形成于山地的草原群落下降到平地,并往东侵移。第四纪时期的冰期与间冰期的交替,促进了南北之间以及山上与山下之间植物的交流,于是逐渐奠定了现代草原的轮廓。青藏高原的草原群落形成最晚,大概在晚更新世(距今2~10万年),才形成目前的景观。随着草原植被的形成也出现了以草原植物为食,生活在草原环境的特定动物类群。
草原生产力与能流
一般说来,由于受到水分条件的限制,草原的初级生产力在所有陆地生态系统中是中等或中等偏下的。据联合国的资料,北美草原地上生物量为每平方米 263~1062克,年净生产量为每平方米 506~1425克。中国草原地上部分生物量,草甸草原约每平方米为200~500克干物质,典型草原每平方米为 75~200克,荒漠草原仅每平方米为 20~100克。中国草原植物年生产量的估计值为每平方米50~1250克,其上限与北美、苏联草原相似,而下限较低,这是因为中国的荒漠草原气候条件比较严酷所致。蒙古荒漠草原生产量的下限为每平方米49.8克,与中国相似。
草原初级生产力与水分条件有密切关系。1964年在非洲的研究发现,稀树草原地上部分净生产量与年降雨量呈线性相关(图5)。
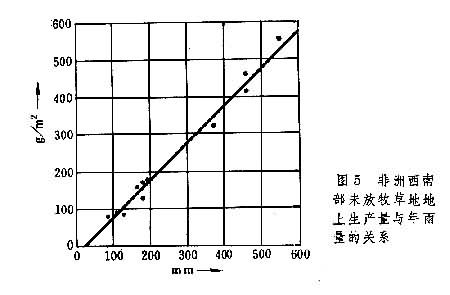
温带草原也存在类似规律。还进一步发现,植物的净光合作用与叶子及土壤的水势呈线性关系。因此,从森林草原至荒漠草原,随雨量的减少,初级生产力有规律地降低。此外,在供水充足的条件下,在年平均气温-2~26℃之间,温度越高则生产力亦越高。因此,稀树草原的生产力显著比温带草原高,平均达1600克/(米2·年),最高可达4000克/(米2·年)。
在生长季草原植物的光能利用率大约为0.1%~1.4%,沿水分梯度而增减。1967年北美高草草原的估算为1.2%,而1979年估算为0.67%~1.09%。联合国在北美洲测定这一数值为0.12%~1.43%。苏联的草甸草原达1.32%,而蒙古荒漠草原尚不及0.1%。
草原初级生产所创造的有机物质为各种植食动物所取食。草原的植食动物多种多样,它们以不同方式采食不同植物以及同种植物的不同部分。植食动物又为肉食动物所食。如鼠被鼬吞食,而鼬被鹰食等等。肉食动物的猎取对象也常常是专化的,捕食者与被食者之间互相制约,组成了复杂的食物网。
草原生态系统中,植物所固定的太阳能量沿食物链传递,逐级减少,所构成的生态金字塔可以美国科罗拉多的矮草草原为代表(图6)。
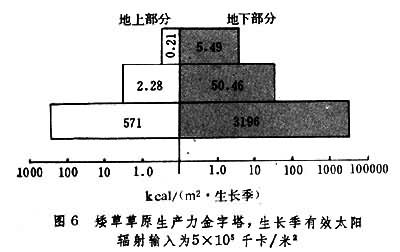
联合国在科罗拉多矮草草原上测定,该地生长季内有效太阳辐射总量为每平方米5×105千卡,绿色植物同期固定的能量为每平方米3767千卡(其中地下部分为每平方米3196千卡,约占总量的85%),生态效率为0.75%;植食动物同期转化的能量为每平方米52.74千卡,生态效率为1.4%(若按草的实际消耗量计算,此数值为11.9%);肉食动物的相应数字为每平方米 5.7千卡与11%。但应注意的是,不同动植物种甚至同一种的不同品系之间,能量转化效率非常不同,选择高光效植物营造人工草地或选用优良畜种,便可大大提高各级转化效率。
在草原生态系统中,大部分能量沿捕食食物链通过,但碎屑食物链也很重要。在原始情况下,草原上凋落物的量相当可观。通常地上凋落物约为地上部分现存量的20%~50%,而地下部分凋落物可能还大于地上部分。在温带草原上,地上枯死部分的分解速率约为两年,地下部分则需 4年左右。凋落物的积累速率大致与分解速率相当,所以在天然草原上常保留着明显的凋落物层。热带稀树草原的分解速率比温带草原快得多,因此,这里一般看不到凋落物层,土壤中也很少腐殖质的积累。
草原生态系统中分解者的生物量相当高,往往超过消费者的生物量。据加拿大南部马特多地区1968年测定数字,该地草原植物量为每平方米434克,30厘米土层内土壤微生物量达每平方米254克;1970年另一次测定,相应数字为每平方米622克与328克。当然在不同地区,这一数值很不相同,北美矮草草原的数值就低得多,如据波尼地区测定,30厘米土层内真菌生物量为每平方米38克,细菌为27.6克,总计为每平方米65.6克(1979)。植物凋落物中约有1/3能量转移给微生物。图7
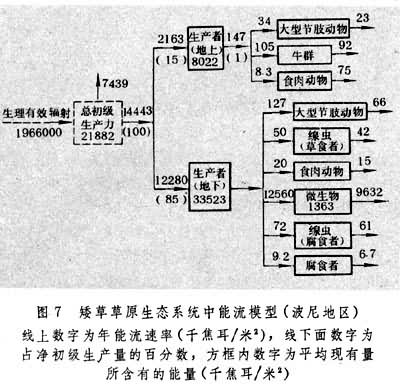
为波尼地区的能流模型。该地区植物生长期内 (154天)的太阳辐射总量为每平方米4158000千焦耳,生理辐射量为每平方米1966000千焦耳,植物净生产量约固定总辐射量的0.34%,生理辐射量的0.73%。植物所固定的能量约15%分配于地上部分,而85%分配于地下部分(在水分条件较好的马特多,相应数字为43%与57%)。植物地上部分能量约有 6.7%流入消费者体内,其余大部分随枯死部分进入凋落物中。植物地下部分的能量大部分转入土壤生物,主要是进入土壤微生物。由于土壤微生物的能量不仅仅得自植物地下部分,因此它的流通总量高于当年流入植物地下部分的总量。
草原的物质循环
物质循环是草原生态系统的另一重要机能。
氮循环
固氮微生物对草原生态系统氮的输入起重要作用,其中共生固氮菌,特别是豆科植物的根瘤菌更为重要。在适宜的生长条件下,根瘤菌在人工草地上的固氮量达每年每公顷100~300千克(1970),高产苜蓿地上甚至可达每年每公顷 670千克(1965)。但多数天然草地上豆科植物较少,如中国及北美草原上豆科植物比例一般不到草群总重量的10%,有时甚至不到1%,因此,这里共生固氮菌的固氮量远低于上述数字。即使如此,它们的固氮量仍是可观的。除豆科植物外,草原上还有一些植物能与根瘤菌共生,如卢多维奇氏蒿、纤细仙人掌等。非共生固氮菌在草原氮素输入中也起一定作用,一般固氮量为1~2千克/(公顷·年);蓝藻的作用不大;而通过降水输入的氮约为3~4千克/(公顷·年)(1980)。大致讲来,植物地上部分氮素周转率至少在50%以上,而地下部分均为19%。研究表明,植物体的氮素只有一部分流入腐殖质,另一部分流入微生物,还有相当一部分以无机氮形式释放出来(ƒ2,ƒ4)。1980年估计,北美矮草草原上从植物体进入土壤腐殖质的氮不超过10%,而腐殖氮的矿化率每年仅为 0.2%。如波尼附近草原上的植物每年需氮量约每年每平方米7克,其中仅1.8克来自腐殖质,5.2克直接来自凋落物。
氮素可以通过动物移动,植物产品的收割、挥发、淋溶等方式流出生态系统而损失。一般情况下,草原上氮的损失量要比农田小得多,因为在干旱、半干旱地区,通过淋溶而损失的氮素微不足道。但在过牧地或施氮肥后,氨态氧可能通过挥发而大量损失。1970年有报道说,排泄在草原干燥土壤上的牛尿中,90%的氨态氮被挥发掉。野生动物的移动对草原养分循环的影响不大,一般家畜摄取养分的80%~90%以粪便等形式返回土壤,它们带走的营养物质通常不超过其摄取量的10%(1956)。总之,在自然状态下氮素的流出量不大。常常是流入量略大于流出量,即土壤库中的氮素逐年积累。但另一方面,系统内氮素的年输入量远不能维持植物年生长对氮素的需求量,如北美矮草草原上植物体所含氮素量达每年每公顷 428千克(1977),这可能是当地氮素年输入量的10倍以上。可见,已存在于系统中的氮素的再循环对草原是重要的。据同一地点测定,0~36厘米土层中的氮素达每公顷3725千克,几为植物体氮素的9倍,这说明参与循环的氮素仅为系统内总氮量的一小部分。
值得指出的是,人为地从草原上取走大量动植物产品,便可使草原的氮素循环失去平衡。例如中国草原区东部的天然割草场,常常由于连年刈割而导致产量、质量持续降低。在这种情况下,则须通过施氮肥或种植豆科牧草以增加草原的氮素输入,保持氮素平衡。开垦原始草原也会使土壤中有机物质的分解加速,收割和水土流失又带走大量的氮。因此,开垦草原实际上是在开采土壤中的氮贮库,因此必须通过增施氮肥来维持氮素平衡。
磷循环
草原土壤中磷的总量以及有机磷对无机磷的比例,与气候条件尤其是气温有密切联系(1961)。如在美国草原,从南往北随着气温的降低,土壤表层15厘米内总磷量呈现有规律的增高。此外,在总磷中可利用的有机磷数量常常超过无机磷,这点与耕作土壤迥异。看来,在草原的磷循环中,有机磷的转移起着重要作用。
植物吸收的磷仅为土壤中可利用磷的一小部分。植食动物所摄取的磷,约有60%以上通过粪便排出,归还土壤,通过动物所移走的磷不到动物吸收总量的10%(1967)。大部分磷在有机物残体中长期保存下来。土壤中有机磷的矿化与有机氮和碳的矿化有密切关系。1977年估计,在北美草原上,磷的矿化速率约为每平方米3~6.4克。分解者对磷的摄取几乎5倍于植物吸收量,有机磷的矿化量与磷的总摄取量接近平衡,因此当生长季结束时土壤中有效无机磷的含量几乎与生长季开始时相等。
草原的开发利用
以单位面积动物量计算,草原在所有陆地生态系统中名列前茅。早在旧石器时代,人们就大量捕杀非洲稀树草原和欧亚大陆草原的野生动物;当人类第一次越过白令海峡移入北美时,那里草原动物也遭到同样的命运。在最近的一次冰期,仅非洲和北美就有61个大型草原哺乳动物的属被消灭,当然这主要由气候的重大变化引起,但人类的捕杀也无疑起了一定作用。当人类学会使用武器之后,摧毁草原野生动物的速度加快了。1870年前后,北美草原上还有千万头野牛到处游荡,随着对美国西部的开发,30余年间已把野牛消灭殆尽。现在,这种动物只能在国家公园内才能见到。欧亚大陆草原上的野马已近绝迹,野驴、野骆驼也已很少。
人类大量捕杀草原野生动物的同时,又把一些对自己有用的动物驯养起来。逐渐地,家养动物代替大型野生动物成为草原上主要的消费者。在原始状态下,草原生态系统中的食物链比较长,食物网也比较复杂。各种动物之间以及动物与植物之间通过相互制约和自然调节而保持稳定与平衡。一旦家畜代替了野生草食动物,种类贫乏了,食物链缩短了,草原生态系统变得比较脆弱。当然,放牧畜牧业较之狩猎是人类社会的一大进步,也给人类提供了更多的动物性产品。但在人口压力日渐增加的今天,不少地区无限制地利用草原,不考虑草原的生产力而盲目增加载畜量,结果使草原生态系统入不敷出,破坏了生态平衡,导致草原生产力的持续降低。因此,在放牧利用条件下,应以草定畜,把牲畜数量保持在能使草原正常更新的范围内,据试验,当牲畜采食量长期超过地上产量的50%时,就会引起产量降低,草质变坏,导致经济收入的降低,只有在这一限度内实行轻牧,报酬才是最高的。因此应在研究不同草原类型的生产力及其动态的基础上制定适宜的载畜量,以保证草原的更新、保持草原的生态平衡。同时,可通过培育牲畜良种以及加快周转等措施,增加单位草原面积的畜产品。
除放牧利用外,今天世界草原的很大一部分已被农田所代替。如北美的高草草原目前已开垦殆尽,成为美国玉米和小麦的主要产区;在匈牙利,昔日的“无林草地”目前几乎皆成为农田;南美东部的草原亦残存无几,大部为农田和人工草地所取代;苏联草原也进行了大面积垦植。在中国,黄土高原早在千年前已被彻底开垦;鄂尔多斯草原也在几百年前开垦殆尽,这些地区开垦之后,由于作物单一和粗放经营,水土流失严重,土壤肥力迅速消耗,不少地段已成为不毛之地,是中国土地单产最低的区域之一。天然草原的土壤质量较高,尤其当禾草中混生适当比例的虫媒传粉的杂类草时,可形成肥力极好的土壤类型,草原黑土与黑钙土便是地球上最肥沃的土壤。草原转为农田,消除了天然禾草与多种杂类草,代之以一年生作物,后者腐殖质形成作用很低,而且其产品被大部分取走。因此,草原开垦后若不通过施肥来补充养分,则土壤肥力很快降低。如中国东北平原的黑土,原始状态下腐殖质层厚达50~70厘米,有机质含量达6%~15%,开荒后4~5年内产量很高,若不施肥,15年后仅及前5年产量的一半,40年后肥力基本耗竭。至于条件较差的草原,这一过程更短得多。可见,草原的开发必须伴随恢复肥力、保持生态平衡的一些措施,否则将导致资源枯竭。目前草原开垦后保持养分平衡的办法有几种:一是轮歇,如美国西部,有些草原开垦后种小麦,呈条带状间隔种植,种一年歇一年,条带间互相轮换。这不但对恢复地力有益,而且对保持土壤水分也起一定作用。二是轮作,作物与苜蓿多年生牧草轮流种植,借助豆科牧草固定氮素,从而保持土地资源的生产力。三是对已垦农田进行灌水、施肥等耕作措施,不断施入补加能量。进行集约经营,保持高的生产力。一般地说,施入的补加能量越多,生产力也越高。但近来不少地区发现,当生产力达到一定水平之后,即使继续投入高额补加能量,生产量的增加也越来越少,而且土壤性状亦趋变坏。因此,利用轮作维持土壤养分平衡的办法日益受到重视。中国草原区的农田,过去开垦后广种薄收,连续单一种植,只取不还,只用不养,结果不少地区土壤肥力及生产力持续下降。今后,草原的利用、改造须考虑到资源的持续利用。在开垦时,就要设计出保持生态平衡的有效措施。
- 参考书目
- 中国植被编辑委员会:《中国植被》,科学出版社,北京,1980。
- A.I.Breymeyer,G.M.van Dyne, Grassland,S ystems Analysis and Man,Cambridge University Press,1980.
- R.T.Coupland, Grassland Ecosystems of the World,Cambridge University Press,1979.
- Helmut Lieth, Primary Production of the Major Vegetation Units of the World.Primary Productivity of the Biosphere, Springer-Verlag New York Inc.,1975.
- Makoto Numata, Ecology of Grasslands and Bamboolands in the World, VEB Gustav Fischer Verlag,Jena,1979.