[拼音]:luanzi chengshu
[外文]:oocyte maturation
卵母细胞成为卵子的过程。即卵母细胞受某些内源因子或外源因子的刺激,经历一系列变化,包括卵表、卵质和卵核(又称生发泡)的变化,成为具有接纳精子、进行雌雄两性原核结合和发育为正常个体等能力的功能卵的过程。在卵子成熟过程中,卵母细胞完成分化,并具备了受精后正常发育所必需的物质和信息贮备。
卵子成熟大致可区分为卵表成熟、卵质成熟和卵核成熟3个过程,它们相互之间有一定的制约关系。以脊椎动物为例,卵表成熟,是指卵表分子的排列趋向规则化,皮质颗粒迁居深层等变化,以确保受精后的正常应答反应;卵质成熟,是指在激素刺激下卵质中出现的系列变化,包括“促成熟因子”(简称MPF)以及与受精后正常发育有关的某些因子的出现;卵核成熟,是指在MPF等的作用下,生发泡破裂,以及破裂后所发生的一系列变化。
卵核成熟之前,卵母细胞都需经过减数分裂使染色体数目减半。过去,卵子成熟是以完成减数分裂为标准,故减数分裂又称成熟分裂。实际上,卵表成熟、卵质成熟和卵核成熟并不同步。在少数动物中,如扁形动物的涡虫和肝蛭的卵母细胞,在卵未长足之前就受精,携带着精子的卵继续生长,而后进行减数分裂,实现两性原核结合,发育为正常个体。在大部分动物中,卵母细胞只有在长足后,经激素刺激,才趋成熟。但是各类动物受精的时刻并不相同,有些动物长足的卵母细胞是在减数分裂Ⅰ前期受精,如马蛔虫卵;有的在减数分裂Ⅰ中期受精,如家蚕卵;有的在减数分裂Ⅱ中期受精,如大部分脊椎动物的卵;有的于雌性原核形成后才受精,如棘皮动物卵。显然,卵母细胞获得受精的能力,并不以卵核成熟为先决条件。
卵子成熟的激素调控
长足的卵母细胞其成熟的起动,决定于促性腺激素调控的卵巢的功能状态。在脊椎动物,促性腺激素由脑垂体合成和分泌,作用于卵母细胞外围的滤泡细胞或颗粒细胞,间接地调控卵母细胞的成熟。促性腺激素对卵母细胞的直接作用尚无法排除。
在促性腺激素的作用下,卵子的成熟过程和排卵过程往往是同步的,但它们之间并无相关性,是两个完全独立的过程。这方面在两栖类和海星卵上的研究最为透彻,在哺乳类卵上的研究也不少。
两栖类卵母细胞周围的滤泡细胞经促性腺激素作用,合成和分泌甾体激素──主要活性成分是孕酮,后者作用于卵母细胞的膜层系统(包括质膜和卵内膜系),诱发成熟。硬骨鱼滤泡细胞分泌的成熟诱导激素为 17α,20β-双羟-4孕烯-3-酮。在海星,促性腺激素刺激卵泡细胞产生1-甲基腺嘌呤(1-me-thyladenine),继而诱发卵子成熟。
卵子成熟的生物化学变化
卵子成熟是一个需能的代谢过程。缺氧情况下,卵子成熟无法起动。随着卵的生发泡破裂,氧耗量逐渐增加,这可能是氧化磷酸化反应强化的结果。
卵母细胞以糖作为能源。乳酸对于深埋于卵泡中而缺氧的卵母细胞是必不可少的。卵母细胞自身缺乏辅酶Ⅰ,不能激活乳酸脱氢酶,必须依赖于周围的滤泡细胞,将乳酸转变为丙酮酸后,才成为卵子可以利用的能源。
在卵子成熟过程中,蛋白质合成普遍增加。如爪蟾卵出现两次蛋白质合成高潮,第1次是在生发泡破裂之前,第 2次是在生发泡破裂之后。蛋白质的合成不仅有量的变化,也有质的变化。不同程度地抑制卵子成熟过程中的蛋白质合成,会不同程度地影响卵子成熟或受精后的卵裂,卵质成熟是依赖于翻译水平上的蛋白质合成。
在成熟早期阶段,卵内大量合成组蛋白并从卵质转移至生发泡中,以供受精后快速分裂的子细胞核使用;这些组蛋白可能起着调控基因活动的作用。
在爪蟾卵母细胞生发泡破裂之前,不仅有rRNA合成,同时也有少量的非均一核糖核酸(hnRNA)的合成;生发泡破裂之后,rRNA合成迅速减少,hnRNA继续合成。尽管卵母细胞的成熟过程中有RNA合成,但卵的成熟并不依赖于RNA合成。
在爪蟾卵母细胞的成熟过程中,卵核内未发现DNA合成活动,在细胞质内少量DNA可能是线粒体合成的。抑制DNA的合成不会影响卵母细胞的成熟。
卵子成熟调控的分子机制
现有的证据表明,在卵母细胞的发生过程中,起调控作用的是一些细胞质因子,有些位于卵的特定区域中,对卵核产生正的或负的效应,改变和决定基因的表达特性。在卵母细胞的分化过程中,细胞质因子仍起着调控作用。其中主要的调控因子是细胞周期蛋白和促成熟因子。活化的促成熟因子能使卵母细胞的生发泡破裂,并引起染色体浓缩、纺锤体形成、第一极体外排、直至停顿在减数分裂Ⅱ中期,同时使卵质具备了促雄性原核形成、两性原核结合和卵裂等能力。
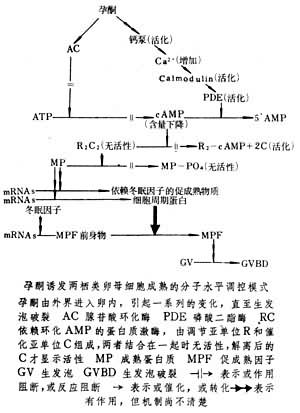
两栖类卵子成熟两栖类某些长足的卵母细胞对孕酮敏感,经孕酮作用于膜层系统,包括卵表质膜和卵内的其他膜层结构,从不同途径使环腺苷酸(cAMP)含量下降(见细胞膜受体)。致使依赖于cAMP的蛋白质激酶的活性降低。在此类蛋白质激酶失活以及在磷酸酯酶-1的作用下,某些蛋白质发生去磷酸化。这些蛋白质的去磷酸化,具有消除抑制和推进成熟过程的作用,故称作“成熟蛋白质”。
孕酮有直接抑制卵母细胞腺苷酸环化酶的作用,也同时有刺激钙泵,使卵内游离钙量增加,从而活化钙调素激活磷酸二酯酶的作用。就这样以抑制cAMP形成和加速cAMP去环化等两条途径,降低卵母细胞内cAMP含量。
由于尚未真正分离到成熟蛋白质,故对于成熟蛋白质进一步导致促成熟因子形成的具体过程,还很少了解。80年代以来,通过对冬眠中华大蟾蜍的卵母细胞和恒定高温饲养条件下中华大蟾蜍的卵母细胞的实验分析,发现冬眠动物卵母细胞质中存在“冬眠因子”,而在孕酮刺激后的高温饲养动物卵母细胞质中出现一种“依赖冬眠因子的促成熟物质”。这种物质不同于促成熟因子,它不能促进高温饲养组动物卵母细胞核成熟,却能在具备冬眠因子的冬眠组卵母细胞中,诱发促成熟因子的出现、促卵核成熟。高温饲养组动物卵母细胞质不具备扩增MPF的作用,因此推测它在孕酮作用下,虽能合成足量的细胞周期蛋白,但因无MPF前身物,致使无法产生MPF。这些发现有助于促进对孕酮诱发成熟的中间过程的认识。
在卵子成熟过程中,促成熟因子的产生并不依赖于卵核或DNA的转录活动,但必须有翻译水平的蛋白质活动参与,以产生一定阈值水平以上的细胞周期蛋白。促成熟因子是一种蛋白质,分子量约为34000,故又简称P34,是裂殖酵母细胞分裂周期基因cdc2编码产物的同类物。促成熟因子具有自我扩增的性能,自身就是蛋白质激酶,能使无活性的促成熟因子前身物磷酸化,显示扩增作用。该扩增过程尚依赖于翻译水平的蛋白质合成活动。
有关促成熟因子导致生发泡破裂,以及破裂后一系列变化的调控机制,尚不清楚,尽管已有报道,MPF能与中心体专一地结合。在生发泡破裂之前或破裂之后,卵质中相继出现一些调控核演变的活性成分,包括能使染色体收缩的染色体聚缩物质,使分裂停滞在中期阶段的细胞分裂静止因子以及具有促精子头部原核化的雄性原核生长因子等(见图)。
 哺乳类卵子成熟
哺乳类卵子成熟
哺乳类卵子成熟的调控机制还不太清楚。在长足的卵泡中,停顿在减数分裂I前期双线期的卵母细胞成熟活动,受到卵泡液内卵母细胞成熟抑制因子(OMI)的制约。OMI是一种小分子量的多肽,来源自颗粒细胞和卵丘细胞。抑制卵母细胞成熟信号的传递,似乎是通过卵丘细胞和卵母细胞之间的间隙连结实现的。促黄体激素(LH)具有解除OMI抑制信号的作用,LH可能抑制OMI的产生,或阻断卵丘细胞和卵母细胞间的信号联系。
除了OMI,其他因素如卵丘细胞与卵母细胞间的间隙连结,cAMP和甾体激素等可能也参与哺乳类卵子成熟过程的调控。一般认为,具有抑制卵母细胞成熟能力的cAMP,是由卵丘细胞产生,并通过间隙连接转运至卵母细胞中;甾体激素在哺乳类卵子上的作用不同于两栖类或鱼类等的情况,没有诱发卵子成熟的作用。一旦哺乳类卵子成熟活动起动,甾体激素(特别雌二醇),似乎具有使卵子抵达第二次成熟分裂中期的保证作用。
卵子与其周围滤泡细胞等结构,各有特点,在正常的生殖季节和正常的生殖环境中,经促性腺激素的作用,它们是能够互相配合、协调、循序前进,达到顺利排卵,成熟、受精和进一步发育的目的。然而,在人工促排卵、促卵子成熟和人工授精时,由于或多或少与自然的情况不同,倘若未掌握好条件,容易使卵子与其围周结构的活动失调,或影响到排卵,或影响到卵子成熟,或影响到卵子的正常受精,或影响受精后的进一步发育。在这一方面,最关键的是要在卵子成熟的恰当时相进行授精。卵子的整个成熟过程,可分为3个时期,即不够成熟时期,适当成熟时期和过分成熟时期。以两栖类或鱼类为例,不够成熟和过分成熟时期的卵子受精后,往往为多精受精,卵子只表现出激动反应,无修整能力(见受精),发育异常,中途夭折;唯有适当成熟时期的卵子受精后才能正常发育。