[拼音]:hesuan de rengong hecheng
[外文]:synthesis of nucleic acid
以核苷或单核苷酸为原料并且不依靠任何天然模板或引物,采用有机合成反应或酶促合成反应进行的寡核苷酸或核酸大分子的合成。
在核酸的理论研究和实际应用上人工合成是一个重要的手段。1957~1965年H.G.科拉纳等人设计并合成了由一种、两种或 3种脱氧核苷酸组成的重复顺序的脱氧寡核苷酸片段,并以此为模板用DNA聚合酶和RNA聚合酶进一步复制和转录,得到了具有对应的互补顺序的长链人工信使核糖核酸(mRNA),再用这种人工mRNA在无细胞体系中进行蛋白质合成。通过分析这样得到的多肽产物的氨基酸顺序和与模板中核苷酸顺序的对应关系破译了遗传密码。
人工合成基因促进了基因工程的发展。如1977年,坂仓等人首先合成了生长激素释放抑制因子的基因,并使之在大肠杆菌中实现了表达,得到了在大肠杆菌中原来并不存在的活性肽(十四肽)。此后,一系列多肽和蛋白质基因,如胰岛素、干扰素和生长激素等的基因相继被合成,并得到了很好的表达,使得这些原来只能从动物组织中得到的含量不多的蛋白质能以细菌发酵的办法大量生产。
1981年中国生化学家王德宝等完成了酵母丙氨酸转移核糖核酸的全合成工作,这是第一个人工合成的具有全部生物活性的RNA分子。
核酸合成包括化学合成和酶促合成两个方面。
化学合成
以核苷或单核苷酸为原料,完全用有机化学方法来合成核酸。由于核苷酸是一个多官能团的化合物,因此,在化学合成中,必须将不希望发生反应的基团保护起来。例如想将图1 中甲、乙两个3′-核苷酸连接起来生成3′-5′-磷酸二酯键,并且要符合甲在乙前的顺序,如果不将某些基团保护起来,势必会发生许多不需要的副反应干扰产物的生成。在这里,只希望核苷酸甲的3′位上的磷酸基团与核苷酸乙的5′位上的羟基反应,因此甲的2′和5′-羟基,乙的2′-羟基和3′磷酸基团都必须保护起来,以排除不必要的副反应。又如图2 所示由核苷酸甲和核苷乙合成二核苷一磷酸的反应式中,R1、R2、R3和R4都是保护基,其作用是使核苷酸甲的3′磷酸基能专一的只同核苷乙的5′羟基反应生成甲-乙顺序的3′-5′-磷酸二酯键。此外,在核苷酸的碱基上如果有氨基(如胞嘧啶、腺嘌呤和鸟嘌呤的环外氨基)也必须加以保护。在DNA合成中,由于脱氧核苷的2′位上是氢而不是羟基,就不必保护。


除了用图2中的两种原料外,也可以将核苷乙改为5′-核苷酸,并把2′和3′位上的羟基保护起来,同2′和5′羟基被保护的核苷甲上的3′羟基反应(图3)。
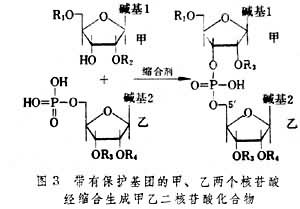
由于这两个反应是通过磷酸单酯和羟基缩合生成3′-5′磷酸二酯,故称为磷酸二酯(合成)法。
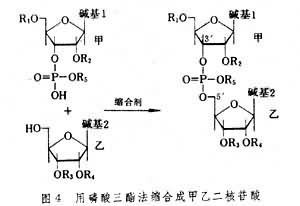
在磷酸二酯法中,由于产物为磷酸二酯,磷酸上所剩下的OH基团,虽然化学活性较小,但也能发生反应,因此,当合成的寡核苷酸链逐渐增长时,由这个磷酸OH基团所带来的副反应也愈益严重,使合成产率急剧下降。目前这个方法已基本上被淘汰,改进的办法之一是采用适当的保护基把磷酸上的OH基团也保护起来,再进行连接反应。即如图4 所表示的那样,用甲核苷3′-磷酸二酯(用R5保护磷酸上的OH基团)同乙核苷的5′-羟基反应生成磷酸三酯型的产物,这个方法称为磷酸三酯法。
另一个改进的办法是采用图5 所示的路线,以保护的亚磷酰胺单体为原料,经1-H-四氮唑活化后与羟基组分连接,先生成亚磷酸三酯,再氧化得到磷酸三酯,称为亚磷酰胺法或亚磷酸三酯法。

在磷酸三酯法和亚磷酸三酯法基础上发展起来的固相合成法,其基本原理是将要合成的寡核苷酸链的3′末端核苷先固定在一个不溶性的高分子上面,然后再从此末端核苷开始,逐步接长,接长的链始终被固定在载体上,过量的未反应物和分解产物则通过过滤或洗涤除去,每接长一个经历一次循环。当整个链的增长达到所需要的长度后,再将寡核苷酸链从固相载体上切落下来并脱去保护基,经过分离纯化得到所需要的最后产物。
固相合成不仅可以缩短合成的时间,提高总产率,并且由于每一个缩合循环都经过同样的操作步骤,因此可以采用自动控制手段。目前已经设计出多种型号的全自动或半自动的合成机器,其每一次的连接产率达到97~99%,最快的每轮循环不到10分钟,最长的合成片段可到100核苷酸以上。
酶促合成
通过酶促反应可以把化学合成的小片段连接成为大片段,或是从已经合成的单链制成双链,它可以加快合成工作的进展,使人工合成核酸大分子的目标得以顺利地实现。
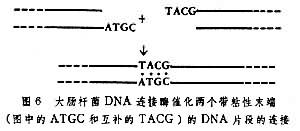
大肠杆菌DNA连接酶和T4噬菌体DNA连接酶是两种经常使用的DNA连接酶。大肠杆菌的DNA连接酶要求两个被连接片段的末端上有几个互补的碱基(称为粘性末端),否则就不能连接(图6)。
T4DNA连接酶不但可以连接带有粘性末端的DNA片段,也可以连接平头的末端(图7)。
已知有几种酶可以用于 RNA合成。多核苷酸磷酸化酶能够以2′位上保护的核苷二磷酸为底物,在一个三核苷二磷酸的引物片段上接长一个核苷酸,以得到的产物作引物,经过同样循环,可以合成更长一些的RNA片段。利用核糖核酸酶N1、T1、U2和牛胰核糖核酸酶A等核酸水解酶的逆反应,在适当条件下,也可以合成某些RNA小片段。T4噬菌体感染大肠杆菌后产生的T4RNA连接酶,是RNA大分子合成中最重要的一个工具酶,它能催化带有5′-磷酸的RNA片段(又称供体)同另一个带有3′-羟基的RNA片段(又称受体)之间的单链连接反应,不仅能够合成RNA小片段,也能合成RNA大片段。
中国科学工作者在人工合成转移核糖核酸的工作中曾经对RNA的酶促合成反应,特别是对有关T4RNA连接酶催化合成RNA的最适反应条件进行过深入的研究,他们在化学合成或化学加酶促合成的 RNA小片段的基础上利用T4RNA连接酶先合成得到6个较大的片段,然后再进一步连接成两个半分子并最后实现酵母丙氨酸转移核糖核酸的全合成。图8是酵母丙氨酸转移核糖核酸的结构和 6个大片段及两个半分子的合成分段位置。
