[拼音]:guhaiyangxue
[外文]:paleoceanography
海洋地质学的一个分支学科,是研究地质时期海洋环境及其演化的科学,又称历史海洋学。它利用现代地质学和海洋学知识,通过海洋沉积物的分析和研究,了解古海洋表层及底层环流的形成、演化及其地质作用,阐明海水成分在地质历史中的变化,浮游和底栖生物的演化,生产力和生物地理发展史及其对沉积作用的影响,以及海洋沉积作用的历史。而古海洋学的中心问题是古海洋环流发展史。
古海洋学与地质学(特别是板块构造说)、古生物学、古气候学的关系密切,它为板块构造理论、生命演化、气候演变提供必要的资料;也为寻找石油天然气、煤、海底锰结核、磷酸盐矿等沉积矿床提供环境指标。因此古海洋学的研究具有重要的实用和理论意义。
古海洋学作为一门独立的学科出现是20世纪70年代以后的事情。1968年开始实施深海钻探计划,许多钻井在各深海大洋区钻穿整个沉积层达到基底。70年代末液压活塞取心技术的出现,能获取大量未扰动的长岩心,为研究中生代以来的海洋环境提供了较为精确的资料。大洋生物地层学,特别是微古生物及超微化石的研究进展迅速,它与磁性地层学、同位素年代学、岩性地层学以及火山灰年代学等技术结合起来,使得全球性地层时代对比和环境分析成为可能。板块构造说为古海洋环境的再造提供了理论基础。古生物学、地球化学及矿物学的现代科学技术广泛应用于古海洋环境的研究,大大推进了古海洋学的发展。现在,利用古海洋学理论和方法,初步建立了中生代以来各时期的海洋古环流、海洋古地理和古气候演变的模式。
研究方法
古海洋学与现代海洋学不同,前者只能利用古生物学、地球化学和沉积学等方法,通过海洋沉积物的分析和研究,间接地确定古海洋各环境要素;后者则采用各种海洋学方法直接测定。
微体古生物法
是古海洋学最主要的研究手段。有孔虫、放射虫、硅藻、颗石藻等微体或超微体生物的生活,主要受海水深度、温度、盐度、浊度、营养盐以及水体运动等各种物理化学条件的控制。这些要素变化的信息便记录在生物个体、生物组合、分异度等特征上,因此海洋生物是海洋环境的灵敏标志。微体古生物方法主要从以下几方面进行分析:
(1)生物时空分布规律的研究。不同生物对其生活环境有一定选择性,如放射虫多见于赤道海域, 硅藻多产于高纬度海区, 窄温性有孔虫, 有的适应于温水, 如截锥圆辐虫(Globorotalia truncatulinoides);有的适应于冷水,如厚壁新方球虫(Neogloboquadriua pachyderma)。根据生物分布还可以推断古海岸线的位置。从底栖有孔虫的居住带和生物分异度可推断古水深。微体生物化石组合和延伸方向大体指明水团和海流流向。窄盐性动植物化石可作为判断海水盐度的指标。生物分异度和生物组合与温度梯度有关,分异度随着纬度升高而降低。海洋动植物的纬向分带标出了气候带。
(2)生物个体形态特征的研究。生物壳体的形态、大小、厚度、密度、旋转方向及骨骼孔隙度等变化,都是为适应生活环境而发生的,它们主要反映水深及水温的变化情况。生物壳体厚度的增减规律与静水压力有关,据此可判明古水深。浮游生物骨骼的孔隙度随着水的密度增大而减小。一些浮游有孔虫壳的旋转方向随温度发生变化,在冷水中多为左旋,暖水中多为右旋,可以用来判别季节变化和气候带。底栖有孔虫的平均寿命在高纬度区比低纬度区大2~3倍。
(3)植物光合作用严格受到海水深度的控制,而海水透光带一般局限在表层200米内,故根据一些植物化石可以判断古海水深度。
地球化学方法
利用海洋沉积物中某些元素和同位素的含量及其比值,可以确定古海水温度、盐度及水团。
同位素指标由于海洋生物壳体的18O/16O比值,一方面随壳体形成时的海水温度而变化,另一方面随当时海水同位素组成(与极地冰盖的消长有关)而变化,因此通过海洋生物化石中氧同位素比值 (18O/16O)的测定,可以计算古海水温度并推断古气候。目前,用于同位素测定的海洋生物主要是有孔虫,此外还有颗石藻、软体动物和珊瑚等化石。海水蒸发作用也能引起氧、碳同位素比值的变化。因此它们也可以用来估算海水盐度。
化学元素指标某些化学元素,特别是痕量及微量元素的溶解度和吸附量随着海水温度和盐度发生变化,也可作为古海水温度和盐度的指标。如方解石中的Mg、Sr的富集随温度而增加,惰性气体Ar、Kr、Xe的溶解度随温度的增加而减小。粘土矿物吸收硼的量与盐度成比例,一般海洋沉积物比淡水沉积物含有更多的硼。海水中的许多痕量及微量元素丰度比淡水中的大,对这些元素进行统计分析可估算古盐度。海洋碳酸盐中的Na和Sr含量大于淡水的,海水的Sr/Ca和Fe/Mn值远大于淡水的。海洋有机质 C/N比是4.3~7.1,反映了以海洋浮游生物(C/N为5~6)为主,而陆源植物的C/N则为30~40。因而它们都是判断古盐度的标志。
沉积学方法
沉积物的矿物、化学成分、结构、构造特征及其空间分布,都可以用于判断古海洋环境。
特征矿物文石或高镁方解石多半出现于浅海环境;而低锶或低镁方解石出现于淡水环境。菱铁矿多半产于10~50米水深的浅海区。自生黄铁矿一般代表海水流动微弱或停滞的还原环境。呈带状分布的石英、高岭石、伊利石等陆源物质和火山灰往往被用来解释海流状况。
岩性特征碳酸盐补偿深度在地质时期历经波动,根据它的波动情况,可以推断古海水深度、生物生产力、水化学特征以及海平面变动情况。蛋白石质二氧化硅、磷酸盐的含量和分布可指示某一时期上升流和生物生产力的状况。深海地层中锰结核的出现,岩性突变、地层和化石带的缺失等,可用来指示沉积间断;而区域沉积间断常可说明地质时期深海底层流及其侵蚀作用的存在。富含有机质的黑色页岩,则可能代表水流微弱或停滞的缺氧还原环境。
粒度和构造特征利用沉积物粒度及其排列方向可以测定古海流的强度和流向,近年来有人试图利用碎屑颗粒长轴导向磁化率来测定古海流方向。有时还可以利用粒度分配特征,大体区分古气候带,例如,以机械风化为主的高纬度区多出现砾石等粗碎屑物质,常伴有分选很差的冰载物质;而以化学风化为主的低纬度带则以泥质沉积为主。根据冰载物质在洋底沉积中的分布状况,尚可用来追索当时洋流的路径与方向。地层中的浊流沉积物往往代表古海洋某种异常事件,如地震、风暴、滑坡等。
初步研究成果
古海洋与古气候演变
深海钻探资料表明,洋底沉积层的年龄不超过 1.7亿年。目前还不可能确切地复原前中生代的海洋演化史。基于深海钻探及其他方面的研究成果,已有可能勾画出晚中生代以来古海洋演化的基本轮廓。古海洋与现代海洋一样,是一个物理、化学、生物和地质作用相互制约的统一体系。晚中生代以来的海洋演化史,贯穿着海水温度由高变低、大洋环流由弱变强的过程,导致晚中生代海洋型式与晚第三纪至现代海洋型式迥然不同。晚中生代海水温度高,极区无冰盖,海洋底层冰水缺乏,温度梯度小,大洋环流弱;晚第三纪至现代则海水温度低,极区有冰盖,出现海洋底层冷水,温度梯度大,大洋环流强。早第三纪海洋作为其间的过渡阶段,它的特点介于两者之间。
大洋环流的演变大洋环流的演变是控制古气候变化(特别是新生代气候变冷)的基本因素之一,而环流的变迁又受到板块运动和陆块间各海水通道开闭的支配。
二叠纪、三叠纪期间(距今约2亿多年前),地球上仅有一个联合古陆,周围是统一的泛大洋(古太平洋),南、北半球大洋中分别存在单一的巨大环流。泛大洋西缘较暖,东缘较冷,南北向的温度梯度反而不及东西向的梯度大。晚三叠世,联合古陆开始解体。侏罗纪北美开始与南美、非洲分裂,形成北大西洋,在闭塞环境中接受了蒸发岩沉积。晚侏罗世至白垩纪,北大西洋东通特提斯海,西南经中美海道连接太平洋,形成了自东向西环绕全球的赤道环流。当时太平洋在南、北半球均发育亚热带反气旋环流和副极地气旋环流。早白垩世南大西洋和印度洋张开,但初期南、北大西洋并不连通,在南大西洋形成大量蒸发岩。1.1~0.9亿年前,南、北大西洋之间出现表层水交流;在0.7亿年前,进一步出现深层水交流。当时北大西洋与北冰洋仍然彼此隔绝,直至新生代初期,北大西洋才开始与北冰洋连通。随着北美大陆西漂与亚洲靠拢,自晚白垩世开始,北冰洋与太平洋之间的深水交流终止。
新生代以来大洋环流的变迁,主要表现在以下三方面:
(1)赤道环流的减弱和中断。(i)4000 万年前印度与亚洲主体汇合,使赤道环流局限于从阿拉伯以北的狭窄水道通过;1800万年前,非洲-阿拉伯与欧亚板块的碰撞和中东造山运动使特提斯海道最终封闭,大西洋-地中海与印度洋之间的交流终止。(ii)新生代晚期,随着澳大利亚北移,印度尼西亚海道关闭,赤道印度洋与赤道太平洋之间的深水交流受阻。(iii)上新世晚期(约350万年前)巴拿马地峡形成,切断了赤道太平洋与赤道大西洋之间的水体交流。
(2)环南极洋流的形成。(i)渐新世最早期,随着澳大利亚与南极洲进一步分离(最初分裂在始新世初),塔斯马尼亚以南的南塔斯曼隆起与南极大陆分开,沿新裂开的塔斯马尼亚海道,印度洋海水流入太平洋。(ii)渐新世晚期或中新世初期,南美洲与南极洲之间的德雷克深水通道张开,完整的环南极洋流最终形成(图1)。
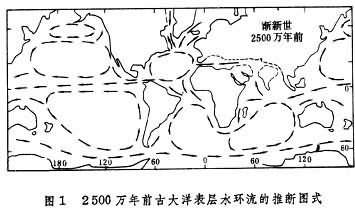
(3)南极底层水的形成。渐新世最早期,随着南极地区冰川和海冰的发展,寒冷的高盐度表层海水下潜,形成南极底层水(冷水圈环流),从而导致大洋底层水富氧,海底侵蚀作用增强,沉积间断及再沉积作用盛行。至晚第三纪,统一的赤道环流已不复存在,环南极洋流十分强劲,开始出现类似于现代的大洋环流特点。
新生代气候变冷过程对深海岩心的氧同位素(18O/16O)分析表明,白垩纪以来气候有变冷趋势(图2)。新生代变冷过程的特点是:高纬地区明显变冷,低纬地区不甚显著;海洋深层水明显变冷,表层水不甚显著;变冷并不是平稳的渐变过程,在温度下降的总趋势上叠加着几次急剧的气候波动。
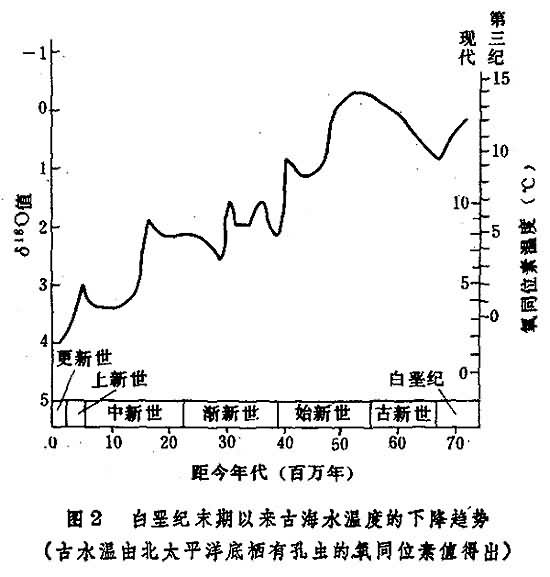
始新世末、中中新世和晚上新世为三次急剧变冷期:
(1)始新世末期事件。距今3700万年前,海洋底层水温骤降4~5°C,南极周围首次形成大规模海冰,开始出现寒冷的南极底层水。南极大陆上已有冰川,但可能尚未成为巨大冰盖。有孔虫等生物蒙受沉重打击,渐新世初期海洋生物的分异度极低。这些情况的发生可能与塔斯马尼亚海道的张开有关。南印度洋高纬海域的表层冷水,经塔斯马尼亚海道注入南极罗斯海域,取代了南下的东澳大利亚暖流,从而触发了南极地区的冰冻。
(2)中中新世南极冰盖形成。大约1400万年前,南极大陆形成冰盖,南大洋海冰进一步扩展(当时北半球仍无冰川),冰载沉积物分布甚广,硅质生物沉积带向北推展。这一时期的海洋进一步变冷与环南极洋流的形成和强化有关,它使南极水体无法与低纬度温暖水体交流,南极大陆在一定程度上处于热隔绝状态。中新世中期冰鸟-法罗海脊沉没,北冰洋-挪威海水注入北大西洋,北大西洋深层水长驱南下,然后上升,加入环南极洋流,这可能为南极大陆带来了形成冰盖所必需的降水。
(3)晚上新世北半球冰盖形成。大约300~240万年前,北大西洋和北太平洋出现冰载沉积物,标志北半球冰盖的形成,由此开始出现冰期-间冰期气候旋回。北半球生成冰盖,可能与巴拿马地峡形成,墨西哥湾流强化有关。湾流为形成冰盖带来了水汽。
第四纪海洋据氧同位素的资料研究,第四纪冰期与间冰期频繁更迭,最突出的变动周期约10万年,其上还叠加着约4万年和2万年的周期。这些周期长短分别与地球轨道偏心率、自转轴倾角及岁差的变动周期相当,因此,冰期-间冰期旋回可由地球轨道变动说(米兰科维奇理论)作出比较合理的解释。随着冰期、间冰期的交替,反复发生海退与海侵,海面变动幅度可达100米左右;深海沉积物的碳酸盐丰度发生周期性变化,构成碳酸盐溶解旋回;气候带(特别是极峰线)南、北迁移,冰载沉积物分布区时进时退。至第四纪后半期,气候变动的幅度达到最大。距今70万年以来北极冰盖增厚。随着温度梯度和大洋环流的增强,南大洋及其他一些海域生物生产力升高,至第四纪晚期达到新生代最高值。在冰期,冰蚀作用强盛,海面下降导致河流侵蚀作用增强,气候干冷导致沙漠扩展,卷扬起大量泥砂,从而使大洋中陆源沉积速率明显增大。
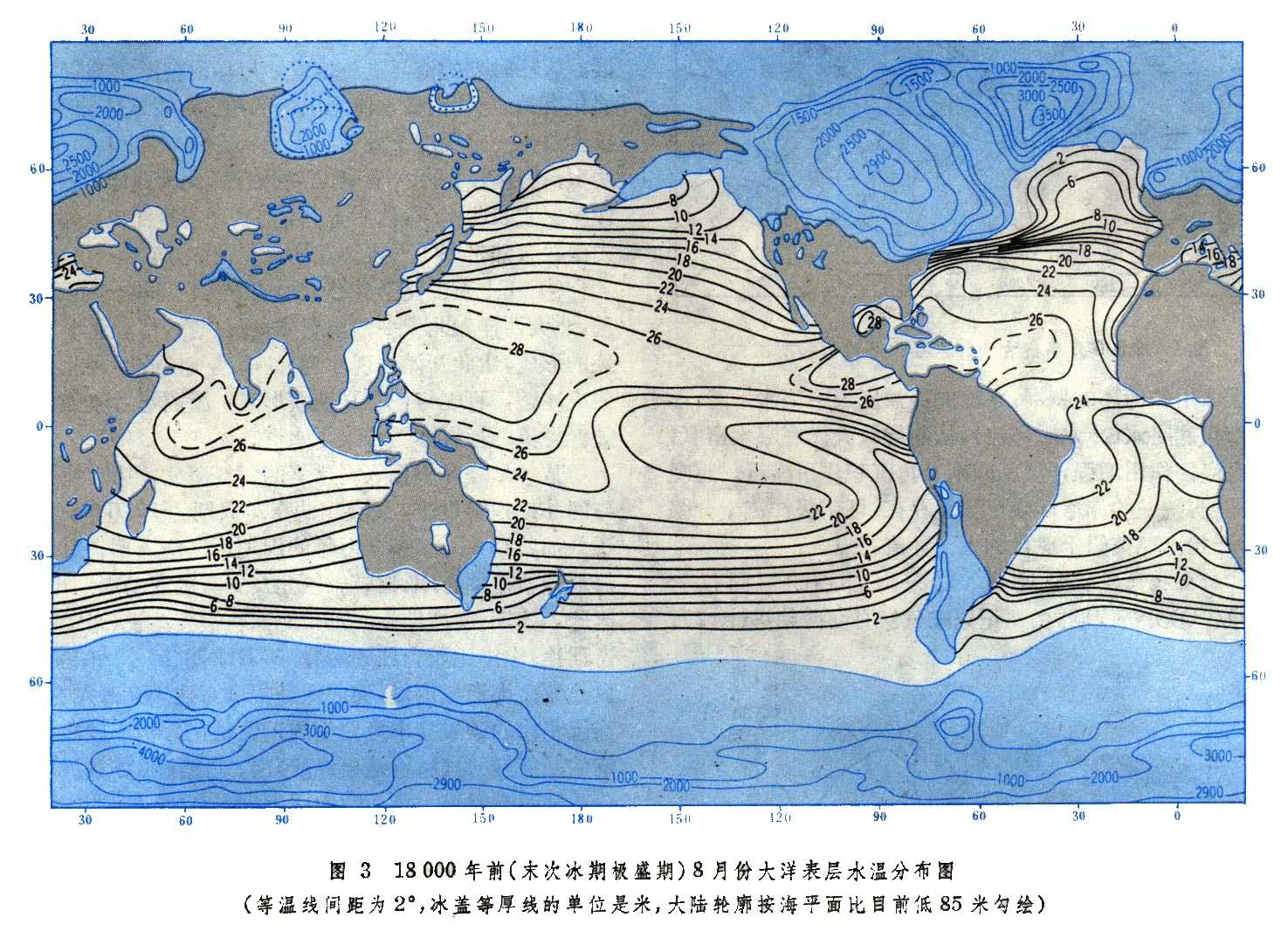
据“远期气候调查、测绘和预报”(CLIMAP)的结果表明:18000年前为末次冰期鼎盛期,当时冰盖广达陆地面积的三分之一,厚可达3公里。海平面至少比现代低85米。18000年前的大洋表层水温分布图(图3)显示:极峰线向赤道偏移,海水温度低,表层海水温度比现在低2~3°C;温度梯度高,尤以北大西洋和南大洋最为显著;沿非洲、澳大利亚和南美洲西岸的东部边界流强盛,伴随着冷水向赤道方向扩展;赤道上升流和沿岸上升流也很活跃。近10000多年来,温度升高,冰川退缩,海面回升。目前地球正处在间冰期。
古海洋演化中的重大事件
深海钻探和同位素地球化学等方面的古海洋学研究,揭示了大洋演化中的一系列重大事件。除上述大洋环流与气候变冷过程中的事件外,重要者有白垩纪中期缺氧事件,白垩纪末生物绝灭事件和中新世末地中海变干事件。
白垩纪中期大洋缺氧事件白垩纪气候暖热而均一,赤道与极地之间以及垂向上的海水温度梯度均小,无底层冷水(由于中、低纬度边缘海强烈蒸发,可能形成热而咸的底层水),大洋环流较弱,致使大洋中缺氧层较宽。缺氧事件的标志是广泛形成纹层状黑色页岩,其有机质含量高达1~30%,缺乏底流侵蚀和生物扰动痕迹,无底栖生物化石。据深海钻探资料,黑色页岩以大西洋分布最广,印度洋和太平洋的无震海岭也有发现;形成时代多在白垩纪中期。当时大西洋可能一度处于缺氧或近于缺氧的停滞状态。但沉积物富含有机质也可能与生物生产力及沉积速率较高有关。
白垩纪末生物绝灭事件距今6600万年以前,恐龙、菊石、浮游有孔虫、超微浮游生物和箭石等大批绝灭。据统计约有二分之一的属消亡了。许靖华等通过深海钻探岩心的研究,发现白垩-第三系界面处,CaCO3含量极低,显示钙质浮游生物数量骤减,碳酸盐补偿深度一度急剧上升;18O/16O比值陡降表明温度突然上升,13C/12C比值聚降,也反映了大洋化学和温度的急剧变化。关于这一事件的起因,众说纷纭。诸如气候变化、海面变动、地磁场倒转、火山爆发等说法,难以解释生物绝灭的突发和全球性质。鉴于界面粘土层中铱含量极高,而陨石中富含铱,一些学者主张星体陨落说。大陨星撞击地球释放出的巨大能量,导致温度突然升高,臭氧层遭到破坏;冲击作用掀起的大量尘埃遮蔽阳光,植物的光合作用受阻,可导致食物链中断而危及一系列生物。
中新世末地中海变干事件深海钻探揭示,地中海海底埋藏着中新世末期(墨西拿期,距今约 500多万年前)的蒸发岩。蒸发岩厚可达2~3公里,总体积100万立方公里,致使大洋水的盐度降低2‰,这一事件使地中海的海洋生物陷于绝境,故称地中海盐度危机,也叫墨西拿事件。1800万年前中东造山运动切断了地中海与印度洋之间的联系,地中海成为西通大西洋的内陆海。中新世末期,由于板块汇聚,地中海西端海峡变浅,加之海面下降,使地中海成为隔绝状态。蒸发岩的上覆和下伏地层均属半远洋沉积,底栖有孔虫的古深度分析表明,地中海在变干前后有类似于现代的深海环境。硬石膏、叠层石和干裂等现象说明这一深海盆地曾经干涸。干涸时期,周围大陆的河谷深切至现代海面以下数百米,河流在裸露的大陆坡上刻蚀出深邃的峡谷。至上新世初期,直布罗陀海峡打开,大西洋水再度进入地中海。
古海洋学是近一、二十年来迅速兴起的新学科,目前还不够成熟,存在不少争论和假说。但作为一门高度综合的学科,它正在将岩石圈、水圈、生物圈和大气圈的研究结合起来,从而有可能为地质科学带来新的重大变革。
- 参考书目
- C.Emiliani,ed.,The Oceanic Lithosphere,The Sea,Vol.7,A Wiley-Interscience Publ.,New York,1981.
- J.P.Kennett, Marine Geology,Prentice-Hill,London,1982.
- T.J.M.Schopf,Paleoceanography,Harvard Univ.Press,Cambridge,1980.