[拼音]:wuxing shengzhi
[外文]:asexual reproduction
又称无配子生殖,即亲体不通过性细胞而产生后代个体的生殖方式,多见于无脊椎动物。脊索动物中的尾索类也进行无性生殖。各式无性生殖的共同特点是参与生成新个体的细胞在遗传物质组成上和亲代的体细胞相同,一般都是含有相同数目染色体的二倍体细胞。无性生殖不经复杂的胚胎发育,也不发生遗传信息的重组,所以子代的遗传物质基础和亲代完全相同。由一个亲体经无性生殖产生的各子代个体都可形成一个无性繁殖系或克隆。无性生殖有利于处在适宜环境下的个体快速增殖,扩大种群数量。在原生动物中,这种生殖仅是以细胞分裂方式产生新个体。多细胞动物由于细胞和组织的分化程度越来越高,其无性生殖必须由成群的细胞或动物身体的特定部位来承担。(见彩图)
分裂
动物身体直接进行分裂,分裂后每一部分都成为一个完整新个体。
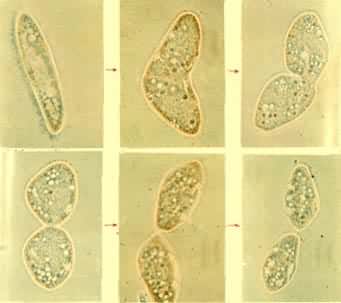
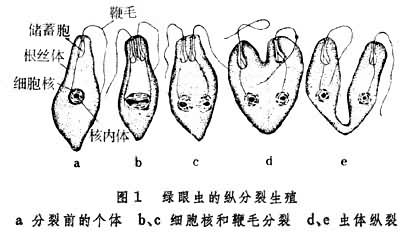
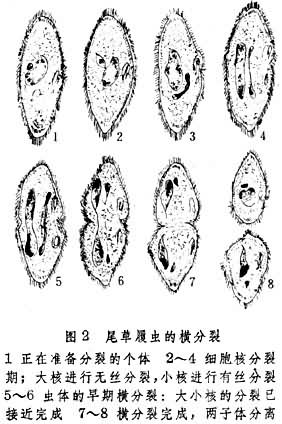
二分裂一个亲体大致均等地分裂为两个新个体,常见于原生动物。分裂面通过虫体长轴时称纵分裂,如绿眼虫 (图1)。分裂面与虫体长轴垂直的则称横分裂,如尾草履虫(图2)。
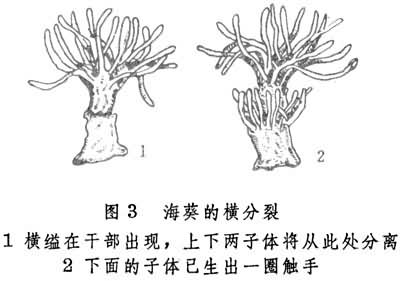
腔肠动物中的海葵也存在横裂式或纵裂式无性生殖。在前种情况下先有横缢出现于茎部,而后虫体由此分为上下两个子体(图3)。
 复裂
复裂
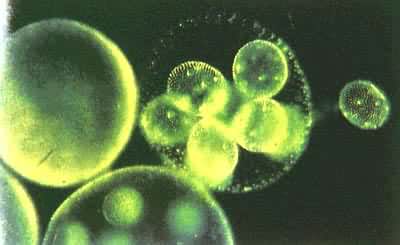
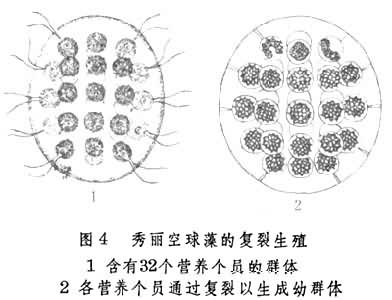
群体鞭毛虫的无性生殖为典型复裂。组成这类动物的营养个员都有一定数目,如实球藻为16个,空球藻为32个。当环境条件适合时,各营养个员同时进行复裂,从而分别发育为与亲体组成相同的新群体(图4)。
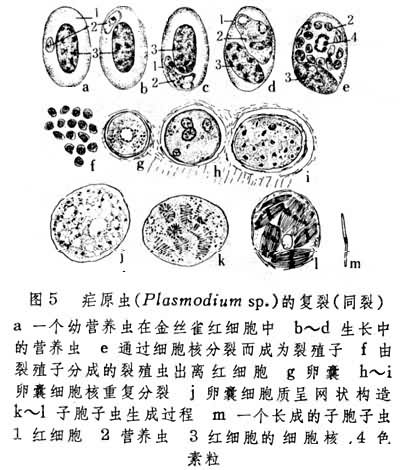
有种较特殊形式的复裂常见于胞子虫类生殖中。其过程是先由亲体细胞核分为若干子核,然后各子核带着一部分细胞质并彼此分离,以形成与其数目相同的新个体。这种复裂又有同裂之称。疟原虫在寄主红细胞中进行的裂殖生殖即属此型复裂(图5)。在此复裂过程中,其核已分而质尚未分的阶段特称为裂殖子。由裂殖子再直接分裂成的许多个体称为裂殖虫。疟原虫受精卵在穿过蚊子胃壁后所形成的卵囊再进行复裂,称孢子生殖,由此形成的大量新个体称子孢子虫。
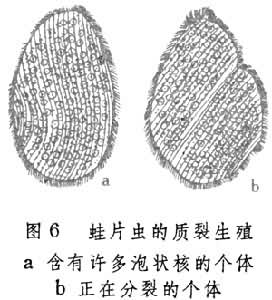
 质裂生殖
质裂生殖
多核原生动物进行这种生殖,在其增殖期间细胞核不分裂,只由细胞质连同原有细胞核分成两个或多个新个体(图6)。
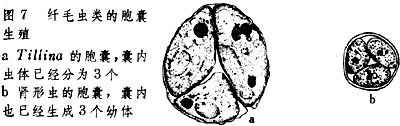
 胞囊生殖
胞囊生殖
有些淡水纤毛虫在环境不适时形成胞囊。其过程包括失去运动细胞器,排出食物胞内含物,由伸缩胞排出体内水分和泌成胞囊等。此种进入休眠期动物待环境转好后,即行分裂(图7):伸缩胞首先在各新生细胞中出现,继而再生出其他胞器,囊膜在酶的作用下溶解,新生幼体释出,发育成新个体。
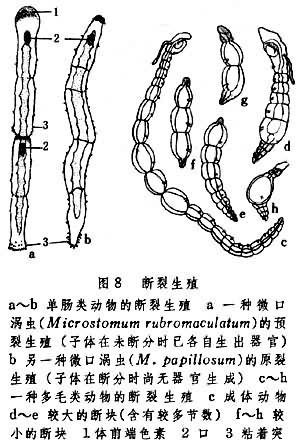
 断裂生殖
断裂生殖
沿动物身体主轴横断为两部或多部,然后由各部分发育成新个体的方式,见于扁形动物中的单肠类和环节动物中的多毛类及寡毛类 (图8)。
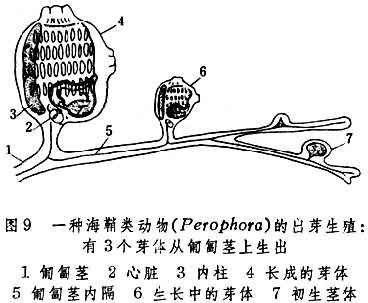
出芽生殖
分外出芽与内出芽两种。
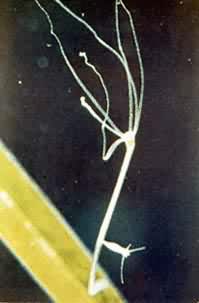
外出芽开始时是从亲体上生出的小突起,后经分化和长大而成为群体中的个员。有的脱离母体,成为独立新个体。原生、多孔、腔肠、蠕虫、苔藓和尾索等类动物都有此种生殖方式(图9)。
 内出芽
内出芽
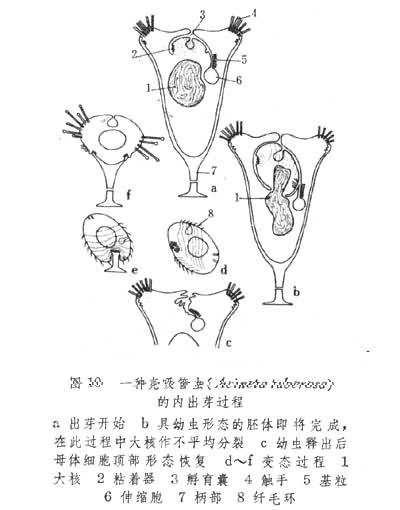
芽体在动物体内生成。其中最简单的是壳吸管虫的内出芽生殖。在此过程中,虫体局部向内凹入以形成胚体。这时亲体中大小核都进行分裂,所生子核进入胚体内。由此胚体长成的具纤毛幼虫脱离母体后,在水中游动,经变态后成为营附着生活的成体(图10)。
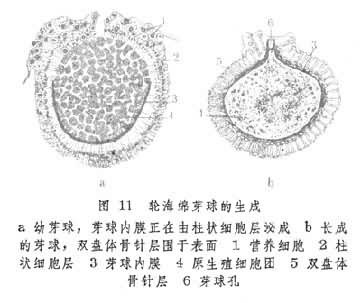
内出芽的另一种方式是芽球萌发,见于淡水海绵中。在环境不适宜的情况下,其中胶层内的一部分原生殖细胞聚成芽球的本体部分。通过吞噬周围营养细胞后这部分细胞内充满营养物质(糖原或脂蛋白)。另有一些变形细胞则特化成柱状细胞层并覆盖在球体周围。由这层细胞向内和向外泌出的物质分别形成芽球内膜和芽球外膜。在两膜之间再由造骨细胞泌成双盘形骨针层(针海绵无此骨针)。此外,两膜间还充以空气。膜上留有一个小孔,称芽球孔。整个坚硬的球形体称为芽球(图11)。淡水海绵在秋天产生大量芽球。母体死后,芽球沉入水底以度过冰冻或干燥的环境。当春天环境适宜时,芽球内的原细胞由芽球孔生出,经过一系列分化和发育过程最后形成新个体。
以休眠芽进行内出芽生殖的情形见于淡水苔藓虫。如羽苔虫休眠芽在虫体的胃绪上生成,开始时是细胞团,以后在表面泌成有保护作用的外壳。虫体死后,休眠芽可抵御不良环境,并可随水漂流,或随风散播到远方,至次年春再发育成新群体。
现存动物中进行无性生殖的只占少数,但这种繁殖方式在动物生殖中有独特作用,因而能在长期进化中得到保留和发展。