[拼音]:guangminsu
[外文]:phytochrome
植物体内的一种蛋白色素。有钝化型和活化型两种形式,分别吸收红光和远红光而相互转化。植物主要通过这种色素接收外界的光信号来调节本身的生长和发育。
发现史
1945年H.A.博思威克、S.B.亨德里克斯和M.W.帕克用分光装置测定了短日植物大豆暗期光间断效应(见光周期现象)的作用光谱,证明这种效应应归因于植物体内的一种吸收红光的色素。以后他们发现红光的光间断效应可以被远红光解除,而再照红光时又可产生光间断效应。红光打破莴苣种子休眠的作用也同样可被远红光可逆地消除。因此设想有一种具有光逆转反应特性的色素存在。1959年W.L.巴特勒等从黄化玉米幼苗体内提取出这种色素,并称之以现名。1964年H.W.西格尔曼和E.M.费热从燕麦幼苗获得了纯光敏素。
光敏素广泛分布于植物界。在被子植物的根、下胚轴、子叶、胚芽鞘、茎、叶柄、叶片、营养芽、花托、花序、发育中的果实和种子中都存在。在蕨类植物、苔藓植物以及某些藻类中也曾观察到光敏素所特有的红光-远红光可逆作用。黄化植株所含光敏素的量比绿色植株高许多倍。但即使在含量最高的黄化幼苗的生长活跃部分(如豌豆上胚轴“钩”和燕麦胚芽鞘节)中的浓度也只在1010-6摩尔/升的水平。在绿色组织中,往往由于高浓度叶绿素的掩盖,而难于检测。
理化性质
光敏素分子由蛋白质和生色团两部分组成。单体(1分子色素和1分子蛋白)的分子量在不同植物中有些不同,变化在 50~150之间。例如燕麦的为60,黑麦的为 120。光敏素的蛋白质中有很大比例的酸性和碱性氨基酸,以及含硫氨基酸,如半胱氨酸。所以光敏素是一个高度带电、非常活泼的蛋白分子,能通过内部重组而改变形式。它的等电点的pH值为6。
光敏素的生色团的每个亚基含有一个开链的四吡咯辅基。生色团以共价键与蛋白质相连。光敏素的两种不同形式(Pr与fr)的吸收光谱不同。Pr(r=red,红)型为蓝色,吸收峰在波长660纳米的红光部分;fr(fr=farred,远红)型为黄绿色,吸收高峰在波长730纳米的远红光部分。两种形式的光敏素吸收相应波长的光后分别转化成另一种形式。由于两种形式的吸收光谱有部分重叠,只是吸收率不同,所以在不同波长的光下进行不同程度的相互转化,形成不同的Pr/fr比值。因Pr对730纳米或更长远红光吸收极少,故在远红光下fr向Pr的转化接近完全;而Pr的吸收峰 660纳米处fr却也有少量吸收,所以在红光下最多只能将80%左右的Pr转化为fr。
每一单体的 fr较 Pr多一个半胱氨酸和一个组氨酸。fr的疏水性较强。
生理作用
在调节植物光形态发生中起作用的是fr,Pr则为生理钝化型。光敏素在调节种子萌发、茎叶生长、下胚轴“钩”的伸直、叶绿体运动、花芽分化以及块茎块根形成中起作用。在种子萌发期和幼苗发育期以及营养生长向生殖生长转化的时期,光敏素的调节作用最为明显。
红光促进莴苣种子等需光种子的萌发。有些种子植物幼苗出土见光后,下胚轴停止生长,下胚轴“钩”伸直,子叶展开。在日光下生长的植物,茎的伸长受抑制;暗期光间断抑制短日植物开花,而促进长日植物开花,这些现象中起作用的都是光下特别是红光下形成的fr。
除了光形态发生效应以外,光敏素还参与调节膜的性质、50多种酶的活动、叶绿素和花青素的合成、乙酰胆碱的含量、合欢等豆科植物的小叶的开合等。此外,光敏素还明显激活rRNA顺反子的转录,加强rRNA前体的形成,以及控制聚核糖体的形成和核糖体的功能。
作用机理
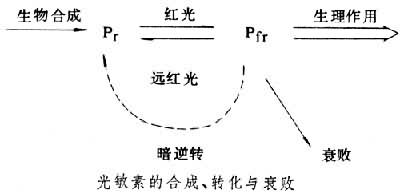
光敏素在体内合成时,Pr形式先形成,在光下一部分转变为fr。在黑暗中生长的植物,如黄化幼苗,只含Pr。fr除在远红光下向 Pr转化外,还会“衰败”,失去活性。此外,在有些植物中,fr在暗中也会转化为Pr,这个过程称为光逆转(见图)。在Pr和fr的相互转化中,还有一系列中间状态,其中有的具有生理活性。在连续光照下,光敏素的各种形式之间达到恒定的比例,Pr合成速度等于fr的衰败速度,光敏素的总量则恒定。离体的光敏素的两种形式也可以在不同波长的光下相互转化,但所达到的Pr/fr比值与同样光下体内的比值不同,说明光敏素在体内的存在状态与离体时不完全相同。由于光敏素的形式和状态的多样性和其间转化速率受各种条件影响的复杂性,使得光敏素在不同植物、不同器官中对各种生长发育过程的调节有许多复杂的情况。光敏素还与植物体内生物钟协同作用,赋予植物以精确计量日照长度的本领。
- 参考书目
- Richard E.Kendrick,Barry Frankland,Phytochromeand Plant Grouth, 2nd ed., Edward Arnold, Ltd.,London,1983.