[拼音]:juelei zhiwu men
[外文]:Pteridophyta
植物界的1门。根、茎、叶中具真正的维管组织,以孢子繁殖。绝大多数在叶片下表面长有孢子囊,并聚集成各式各样的斑点或线条状的孢子囊群,初时为绿色,老时为锈黄色,有的裸露,有的具各种形状的盖。本门植物又称羊齿植物。约有71科381属12000种。广布世界各地,尤以热带、亚热带为最丰富。中国有63科 224属约2400种。蕨类植物不开花结果,一般从外形上难以和种子植物相区别。它形体多样,从高不到 5毫米的微小草本,到高可达20米的乔木状植物。在生态习性上,有水生、土生、石生、附生或缠绕树干。叶片从单一到各式各样的复杂分裂。
形态发生
配子体
有性世代的简单植物体,由孢子萌发而来。孢子萌发后形成几个细胞的简单丝状体,经过顶细胞和侧生分生组织的细胞分裂形成片状体,或称细胞片,再发育为成熟的通常背腹扁平的心形原叶体,原叶体靠近尖端下面生有假根,借以固定和吸收水分,凹缺处为生长点,雌器(颈卵器 archegonium)生长在附近,其外形大而似瓶状,腹部埋在组织内,颈部弯向雄器所在部位,口部突出。雄器(精子器antheridium)呈突起状或圆球形,有柄或无柄,生长在假根附近,通常它的壁由 3个细胞:茎细胞、环细胞和帽细胞(如果帽细胞分裂,则为四个)组成,进化类型的由更多的细胞组成。每个颈卵器含一雌配子──卵子,每个精子器含多个雄配子──精子,精子顶部有纤毛。配子体这样的结构,巧妙地利用了贴近地面,易于获得水分的特点。通常雌配子器的发育晚于雄配子,成熟后除卵细胞外,颈卵器内其他细胞被分泌的有机酸溶解变成胶质而溢出。当原叶体下面有一层薄水层时,精子器的帽脱开(进化类型的为孔裂),精子被释放,通过纤毛运动和受颈卵器口溢出胶质的化学感应而被吸引到颈卵器中,和卵子结合而产生合子,合子萌发后形成胚,再发育成常见的孢子体(图1)。
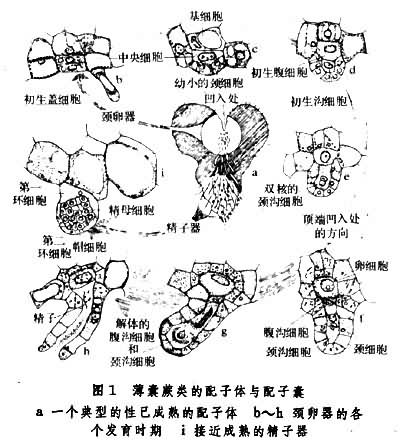
配子体通常寿命很短,在孢子长出后不久即死去,这是由于原叶体结构简单,无维管组织。但也有生活期长的,如翠蕨 Anogramma,在孢子体长出几片叶后,原叶体仍然可见。有些种的配子体可生活4~5年。这些生活期长的配子体,通常较大而厚,出现在较原始的类群。
孢子体
无性世代植物体,具根、茎、叶和输导系统的分化。
(1)根:蕨类植物只有不定根而无主根,通常丛生于直立的根状茎末端,或散生在横走根状茎腹面,借以固定根状茎。根内具原生中柱,无次生生长,不能加粗,表面生有根毛,吸收水分和营养物质。
(2)茎:除桫椤科和蚌壳蕨科的部分种类具高大乔木状直立茎外,绝大多数蕨类植物的茎都贴近地面称根状茎,或短而直立,斜升,或细长横走,少数为缠绕藤本,极少数呈灌木状。其顶端通常被毛或鳞片,有的种类还被叶柄残茎组成的防护罩所保护。
从横切面看,茎的表皮层由外层加厚细胞组成,其内为皮层及中柱体。蕨类植物的中柱体较为复杂,随科属而不同,有时同一株植物随不同发育年龄而有所改变,如桫椤的茎,其基部为原生中柱,中部为管状中柱,顶部为网状中柱。中柱的主要类型有:原生中柱,中柱中央分化为木质部,内层为韧皮部;管状中柱,中柱中央为薄壁细胞组成的髓,向外为木质部和韧皮部,由于韧皮部的位置不同,又可分为外韧管状中柱和双韧管状中柱,前者仅木质部外围有一圈韧皮部,后者木质部的内外均有一圈韧皮部;网状中柱,由管状中柱分裂而成的一种,但维管束仍呈管状排列;多环中柱,网状中柱的各个维管束再次分裂成不规则的分散状排列。
和种子植物一样,蕨类植物的茎的维管束由管胞、筛管、薄壁组织和内皮层组成,但无形成层,不能进行次生生长。在高大如乔木的种类中,其茎干也只是由于薄壁细胞的增加而有限的加粗。
(3)叶:通常兼有营养和生殖两种功能,即绿色的叶片在全部或部分羽片的下面一定部位,都可产生孢子囊,是谓叶片一型。有的科属在同一植株上,先后长出两种不同形状的叶片,一种为正常的绿色营养叶或称不育叶,另一种幼时为绿色,不久即长出孢子囊而失去绿色的孢子叶,又称能育叶,这种类型叫叶片二型。少数种类,如紫萁属 (Osmunda)中,同一片叶上生有能育和不育两种不同形状的羽片,这叫羽片二型。
传统上根据叶片不同把蕨类植物分为两大类:小叶型蕨类,或称拟蕨类,叶小,长宽仅数毫米(如石松、卷柏),或退化成鳞片状(如松叶蕨),或为膜质(如木贼),仅水韭叶大而成线形,但和其他拟蕨一样,无柄,都只有一条中肋而无完善的叶脉系统(据最新发现,热带的石松中有个别种具简单的分枝叶脉),孢子囊生于叶腋;大叶型蕨类,即真蕨类,除槐叶的沉水叶变为须根状,少数属种的叶轴顶端延伸成鞭伏,能着地生根,以及部分科的植物叶轴能无限生长外,其余都为正常叶。幼时通常拳卷,长大分叶柄和叶片两部分,除单叶外,复叶的叶片有叶轴。按分裂的次数不同,而有一回、二回……羽状复叶;羽片,一回小羽片、二回小羽片……;以及羽轴、一回小羽轴、二回小羽轴……之分(图2),在一回羽状复叶中,如果羽片边缘分裂,则按分裂的深浅,称之为叶片一回羽状,羽片浅裂,或叶片一回羽状,羽片深裂(达羽轴的狭翅或阔翅),习惯上也统称之为二回羽状浅裂,或二回羽状深裂(达羽轴的狭翅或阔翅)。如果羽片裂达羽轴,裂片彼此分离,或形成小羽柄,则称这种叶片为二回羽状,其分离的裂片称小羽片。依此类推,直至末回裂片。

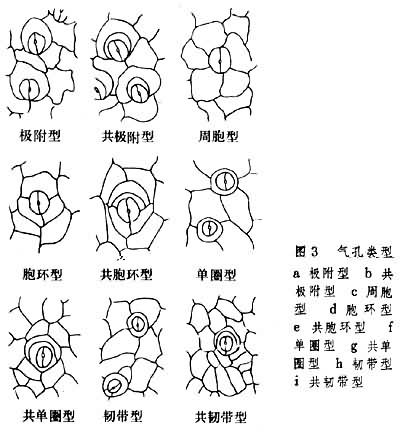
蕨类植物的叶片结构除少数属种有栅状组织和海绵组织分化外,一般仅有海绵状、多少有空隙的叶肉层,有的甚至连叶肉层也没有,光合作用全靠表皮来完成。通常上表皮不具气孔或有极少气孔,下表皮的气孔类型很多,因属种而不同。但也有同一种叶片下表面具好几种气孔型的(图3)。
蕨类植物的叶脉比较简单,绝大多数是分离的,少有简单网结,网眼无内藏小脉或仅有少数内藏小脉(图4)。

有些属种,叶片侧脉的分出和小羽片的分出一样有上先出和下先出之分。先从各回羽轴上侧分出的小羽片,叫小羽片上先出,反之叫下先出。同样,先从主脉上侧分出的侧脉叫上先出脉,又叫上行脉序,反之叫下行脉序。这一特征,在某些属种中稳定而有鉴别价值。
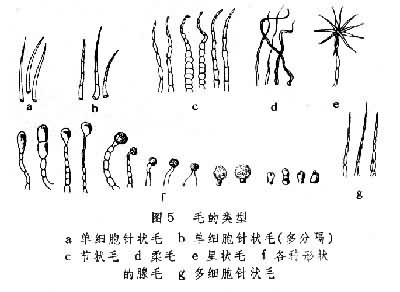
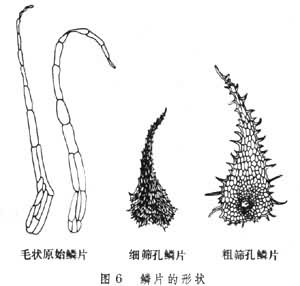
(4)毛被:蕨类植物的毛被相当复杂,分毛和鳞片两大类,或调节叶面空气湿度,或保护孢子囊群。有些腺毛可能和代谢有关,分泌蜡质粉末的腺毛,有保持水分的功能(图5、图6)。
(5)孢子囊:无性繁殖器官,产生在叶片的一定部位,由于它的构造和形成的不同,可分为厚囊和薄囊两大类,前者的囊壳由多层细胞组成,后者的囊壳仅 1层。在发育上,两者的原始细胞都经横断分裂成上下两枚细胞,所不同的是厚囊蕨类的上枚为囊壳细胞,下枚为孢原细胞,而薄囊蕨类的下枚细胞为柄细胞,以后发育成孢子囊柄,上枚细胞经鱼鳞式斜分裂成四面体,再分裂为囊壳和孢原细胞。因此厚囊蕨的孢子囊壁厚而无柄,薄囊蕨的孢子囊壁薄而有柄(图7)。
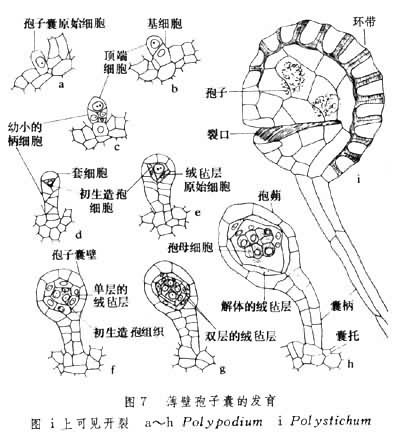
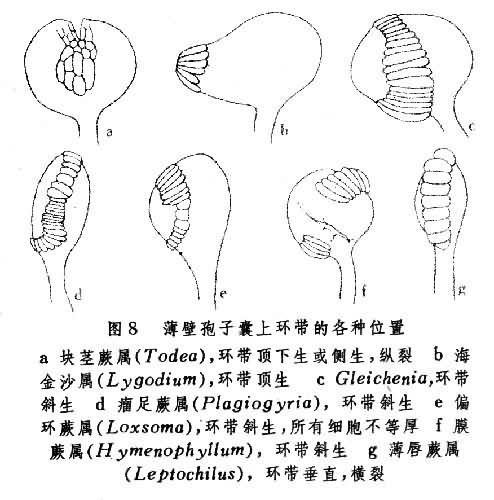
(6)环带:薄囊蕨类孢子囊释放孢子的机构。由数个到数十个 U形加厚细胞和多个扁平的薄壁细胞(包括唇细胞)组成,环绕囊壳。当孢子成熟时,由于环带的U形细胞失水收缩而产生的拉力,孢子囊的唇细胞被拉开,而将孢子弹出。环带的有无和存在部位,常和进化有一定关系。通常较原始的无环带,如厚囊蕨,或仅有不发达的环带(如紫萁),逐渐发展为顶生环带(如海金沙)、横生中部(如芒萁)、斜行环带(如瘤足蕨)和进化类型的纵型而中断于囊柄的水龙骨型环带(图8)。
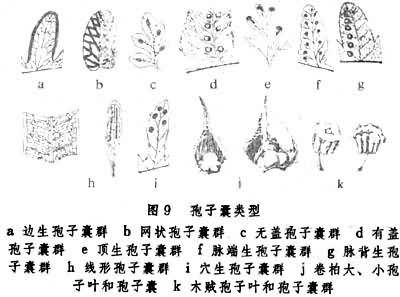
(7)孢子囊群:拟蕨类的孢子囊常近轴而单生于叶腋,其中有些属种,如石松、卷柏的一些种和木贼属。长孢子叶的枝缩短而形成或长或短的孢子囊穗。真蕨类孢子囊通常远轴而生于叶片下面(原始类群的生于叶缘)。它们通常多数聚生在位于叶脉的表皮分化形成的囊托上,而呈各种形状的孢子囊群 (sorus),有些厚囊蕨类的孢子叶常形成穗状或圆锥花序状。较原始类群的孢子囊群的孢子囊数目少而同时发育,发展为从内向外逐渐发育,而进化类型的孢子囊群中的孢子囊为混合发育,即一个囊群里有的发育早,有的发育晚或败育,这在有些种里,败育的孢子囊就成为一种隔丝(图9)。
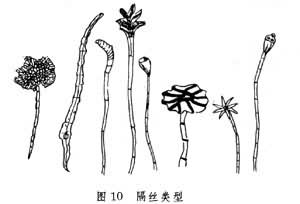
(8)隔丝:又称夹丝。是一种毛状不育器官,混在孢子囊群中起内外保护作用。一般都有长柄,顶部呈各种形状,覆盖幼孢子囊群,如石韦的星状毛隔丝,瓦韦的盾状隔丝等。也有许多隔丝。虽有长柄,但不伸出囊群之外,夹在囊群中起内保护作用(图10)。
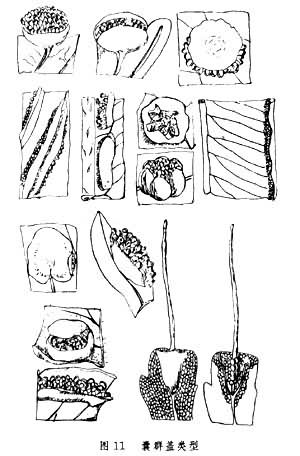
(9)囊群盖:在薄囊蕨纲中异型孢类的孢子囊有一个由叶片特化形成的外壳,包被在大、小孢子囊外面,称之为大、小孢子果,而同型孢类的孢子囊群大都有囊群盖。它主要由叶片表皮细胞分化而来,初时绿色,老时黄褐色,用以保护孢子囊群。囊群盖由于在囊托上形成的位置不同而有上位和下位之分。囊群盖环绕囊托的基部生出,把孢子囊包在盖内的叫囊群盖下位,或叫上位囊群,而囊群盖生于托的顶端或一侧,从上面覆盖孢子囊群的叫囊群盖上位,有的虽生于一侧,但被成熟的孢子囊压在下面,有如半下位。还有些孢子囊群靠近叶边生的,其叶边往往多少特化变质而向下反卷覆盖孢子囊群,这种称为假盖。而有些种类的盖则以毛或鳞片形式出现(如岩蕨属中),囊群盖的形式多样,为鉴别属的重要特征(图11)。
生活史
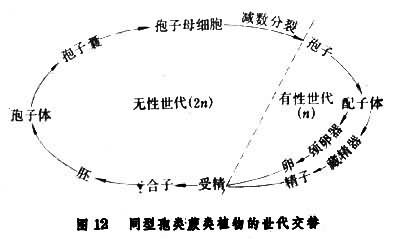
通常肉眼所见到的绿色蕨类植物即是它的孢子体,孢子体上产生孢子囊每一个孢子囊内产生大量的、肉眼看不清的、直径 10~100微米的孢子。孢子比重小,悬浮于空气中,随气流或水流漂移到各处,在适当的环境中萌发,长出微水的绿色(少数不具叶绿素,不能自养)叶状体──配子体(原叶体)。除部分拟蕨类和水生真蕨类孢子体产生大、小两种孢子,萌发后成长的配子体有雌雄之分外,绝大多数蕨类都属于同型孢类,它们的孢子无大小之分,并在孢子萌发后产生的配子体上的不同部位,产生颈卵器和藏精器。并由此分别产生雌配子──卵子和雄配子──精子,雌雄配子结合后形成合子,然后发育成胚,再成长为常见的绿色孢子体。这个过程就是蕨类植物特有的世代交替生活史(图12)。在整个生活史中,常因环境等条件的影响而改变其中某一过程,导致一些不正规的繁殖。常见的为无配子生殖和无孢子生殖。
无配子生殖
生活史中,配子体不产生配子而直接长出新孢子体。但这种配子体或是由仅经过有丝分裂产生的2n孢子萌发而来,或是孢子体先产生复二倍体,再经过减数分裂产生2n的孢子萌发而来(图13)。这种现象在热带产的同型孢类中和石松的一些种中很常见。

无孢子生殖
在孢子体上直接产生配子体,在自然界这是多倍体形成的一条途径。在实验条件下,控制培养基的成分和比例常可诱导出这两种现象。有人认为这和能量有关,能量充裕出现前者,而能量贫乏则出现后者。
此外,在自然界营养繁殖的现象也很普遍。在孢子体的一些部位(如叶轴、羽轴或叶脉)能产生芽孢。还有的叶轴顶端常延伸成鞭状,能着地生根,产生新株。
蕨类植物的孢子体和种子植物一样具根、茎、叶和输导系统的分化,能适应陆地生活,获得更多的营养,从而产生和种子一样起生殖和传播作用的孢子。虽然孢子没有种子在结构机能上的许多进步特征,但由于它的量大,体积小,能被风吹到很远地方,总有一些会遇到适宜的环境而萌发,因此,在某种意义上说更便于繁殖。
种子植物生活史的有性过程是在子房中度过的,而蕨类植物则和苔藓植物一样,配子体能独立生活,也就是说有明显的世代交替。所不同的是苔藓植物是配子体世代占优势,孢子体寄生在配子体上;而蕨类植物是孢子体占优势,分别独立生活。这一生活史特点表明蕨类植物是介于苔藓和种子植物之间的一群植物。它的形态结构决定了它既是高等孢子植物或颈卵器植物,又是原始的维管束植物。
分类及系统
1940年中国蕨类植物学家秦仁昌将经典的十分混杂的水龙骨科(实际上是薄囊蕨纲的绝大部分)划分为33科,促进了对蕨类植物系统的广泛探讨。世界上现有蕨类分类系统不下10个,大体上有两类:其一是以美国E.B.科普兰(1947)为代表的系统,其次是秦仁昌 (1940,1954,1957,1978)为代表的系统(见表)。各国学者根据各自所掌握的材料和观点而有所增减或变化,总的说来,在绝大多数属的划分上,国际蕨类植物学界分歧性不大,而在科,特别是科以上等级排列还莫衷一是。1940年秦仁昌在将水龙骨科分为33科的同时,提出了 5条进化干,即双扇蕨类、桫椤类、蚌壳蕨类和两个绝灭祖先类,表示出各干上科的关系。其后许多学者提出系统树的设想,但由于这些系统树各自根据单方面材料,如印度学者B.K.奈亚尔等主要根据形态解剖方面材料,美国W.H.小瓦格纳等主要根据孢子囊方面材料,意大利的 R.E.G.皮基-塞尔莫利则偏重于染色体的材料等而过于简单化,有的虽有综合考虑,但例证不多,特别是目前在化石证据不足,生物系统学研究也还没有普遍开展的情况下要提出一个比较自然的系统树,还有待于全世界的共同努力。
