[拼音]:yousi fenlie
[外文]:mitosis
真核生物细胞分裂的基本形式。也称间接分裂或核分裂。在这种分裂过程中出现由许多纺锤丝构成的纺锤体,染色质集缩成棒状的染色体。1882年W.弗勒明最先将此种分裂方式命名为有丝分裂。(见彩图)
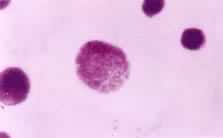
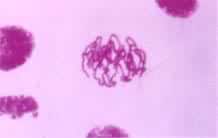
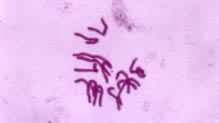
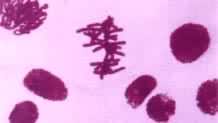


通过有丝分裂,作为遗传物质的脱氧核糖核酸(DNA)得以准确地在细胞世代间相传。通过有丝分裂和细胞分化才能实现组织发生和个体发育。致癌因子通过使有丝分裂的调控失常而导致肿瘤发生。
细胞在进行有丝分裂时,核和胞质都发生形态上的变化,称有丝分裂期。两次有丝分裂之间称为分裂间期。有丝分裂期和间期合起来称细胞周期。有丝分裂的准备工作,例如DNA复制、组蛋白合成、微管蛋白合成、能量准备等都是在间期进行的。
动植物的许多组织细胞都进行有丝分裂。特别是胚胎发育时期细胞的有丝分裂旺盛。高等植物的根尖和茎端分生组织,高等动物的骨髓和一些上皮组织,都是观察有丝分裂现象的适宜部位。
过程
有丝分裂是一个连续的过程,为了描述方便起见,习惯上按先后顺序划分为前期、中期、后期和末期四个时期,在前期和中期之间有时还划分出一个前中期。
前期自分裂期开始到核膜解体为止的时期。间期细胞进入有丝分裂前期时,核的体积增大,由染色质构成的细染色线逐渐缩短变粗,形成染色体。因为染色体在间期中已经复制,所以每条染色体由两条染色单体组成。核仁在前期的后半渐渐消失。在前期末核膜破裂,于是染色体散于细胞质中。动物细胞有丝分裂前期时靠近核膜有两个中心体。每个中心体由一对中心粒和围绕它们的亮域,称为中心质或中心球所组成。由中心体放射出星体丝,即放射状微管。带有星体丝的两个中心体逐渐分开,移向相对的两极 (图1)。这种分开过程推测是由于两个中心体之间的星体丝微管相互作用,更快地增长,结果把两个中心体(两对中心粒)推向两极,而于核膜破裂后终于形成两极之间的纺锤体。
 前中期
前中期
自核膜破裂起到染色体排列在赤道面上为止。核膜的断片残留于细胞质中,与内质网不易区别,在纺锤体的周围有时可以看到它们。
前中期的主要过程是纺锤体的最终形成和染色体向赤道面的运动。纺锤体有两种类型:一为有星纺锤体,即两极各有一个以一对中心粒为核心的星体,见于绝大多数动物细胞和某些低等植物细胞。一为无星纺锤体。两极无星体,见于高等植物细胞(图2)。
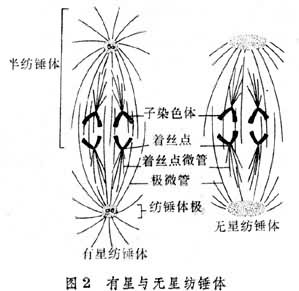
曾经认为有星纺锤体含有三种纺锤丝,即三种微管。一种是星体微管,由星体散射出的微管;二是极微管,是由两极分别向相对一级方向伸展的微管,在赤道区来自两极的极微管互相重叠。现在认为极微管可能是由星体微管伸长形成的。三是着丝点微管,与着丝点联结的微管,亦称着丝点丝或牵引丝。着丝点是在染色体的着丝粒的两侧发育出的结构。有报告说着丝点有使微管蛋白聚合成微管的功能。无星纺锤体只有极微管与着丝点微管。
核膜破裂后染色体分散于细胞质中。每条染色体的两条染色单体其着丝点分别通过着丝点与两极相连。由于极微管和着丝微管之间的相互作用,染色体向赤道面运动。最后各种力达到平衡,染色体乃排列到赤道面上。
中期从染色体排列到赤道面上,到它们的染色单体开始分向两极之间,这段时间称为中期。有时把前中期也包括在中期之内。中期染色体在赤道面形成所谓赤道板。从一端观察可见这些染色体在赤道面呈放射状排列,这时它们不是静止不动的,而是处于不断摆动的状态。中期染色体浓缩变粗,显示出该物种所特有的数目和形态。因此有丝分裂中期适于做染色体的形态、结构和数目的研究,适于核型分析。
后期每条染色体的两条姊妹染色单体分开并移向两极的时期。分开的染色体称为子染色体。子染色体到达两极时后期结束。染色单体的分开常从着丝点处开始,然后两个染色单体的臂逐渐分开。当它们完全分开后就向相对的两极移动。这种移动的速度依细胞种类而异,大体上在0.2~5微米/分之间。 平均速度为1微米/分。同一细胞内的各条染色体都差不多以同样速度同步地移向两极。子染色体向两极的移动是靠纺锤体的活动实现的。
末期从子染色体到达两极开始至形成两个子细胞为止称为末期。此期的主要过程是子核的形成和细胞体的分裂。子核的形成大体上是经历一个与前期相反的过程。到达两极的子染色体首先解螺旋而轮廓消失,全部子染色体构成一个大染色质块,在其周围集合核膜成分,融合而形成子核的核膜,随着子细胞核的重新组成,核内出现核仁。核仁的形成与特定染色体上的核仁组织区的活动有关。
细胞体的分裂称胞质分裂。动物和某些低等植物细胞的胞质分裂是以缢束或起沟的方式完成的。缢束的动力一般推测是由于赤道区的细胞质周边的微丝收缩的结果。微丝的紧缩使细胞在此区域产生缢束,缢束逐渐加深使细胞体最后一分为二。
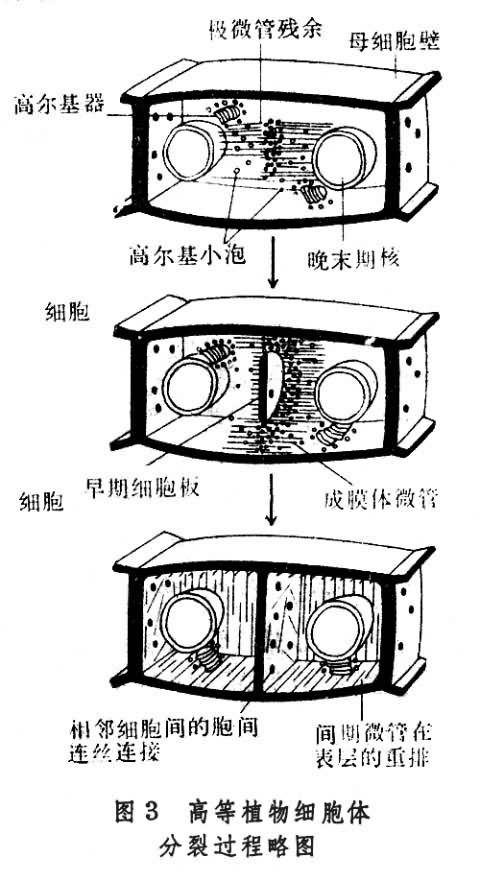
高等植物细胞的胞质分裂是靠细胞板的形成。在末期,纺锤丝首先在靠近两极处解体消失,但中间区的纺锤丝保留下来,并且微管增加数量,向周围扩展,形成桶状结构,称为成膜体。与形成成膜体的同时,来自内质网和高尔基器的一些小泡和颗粒成分被运输到赤道区,它们经过改组融合而参加细胞板的形成。细胞板逐渐扩展到原来的细胞壁乃把细胞质一分为二(图3)。细胞板由两层薄膜组成,两层薄膜之间积累果胶质,发育成胞间层,两侧的薄膜积累纤维素,各自发育成子细胞的初生壁。
机制
染色体的集缩构成染色体的细线在分裂前期缩短变粗,染色体的这种集缩运动是通过染色线的螺旋化实现的。染色质浓缩过程和细胞质中的某些因素有关。如果用实验方法使分裂期细胞与间期细胞融合,可以观察到间期细胞染色质会提前集缩成染色体。这说明分裂期细胞的细胞质中有某种物质能促使染色体集缩。
纺锤体的形成由微管蛋白聚合成纺锤体微管的过程。微管蛋白的聚合有两种基本形式:一种是自我装配型,另一种是位点起始装配型,后者有特殊位点做为聚合的起始部位,前者没有这种特殊位点。形成纺锤体时的位点统称为“微管组织中心”(MTOC)。中心体和着丝点都是MTOC,它们在离体情况下都能表现出使微管蛋白聚合成微管的能力。纺锤体的形成显然和这些MTOC的活动是分不开的。
中期染色体运动用药物(秋水仙素、巯基乙醇等)破坏纺锤体,则染色体不能排列到赤道面,除去药物后,纺锤体重新形成,则染色体又能排列到赤道面,由此可见,染色体向赤道面的排列和纺锤体的活动有关。由辐射损伤或其他原因造成的没有着丝点的染色体断片不能排列到赤道面上。因此说明,染色体向赤道面的排列和着丝点的活动有关。用微束紫外线照射减数分裂时二价体的一侧着丝点或着丝点丝,则染色体不能正好位于赤道面,而偏近于未受照射的着丝点所面向的一极。这说明染色体在赤道面的配位必须两个着丝点及与两极相连的两侧着丝点丝都正常地发挥作用。根据以上事实和其他观察,推测在前中期时两个着丝点分别以着丝点丝与两极相连,靠两极牵引力的平衡,使染色体位于赤道面上。除这种牵引平衡的力量外,还可能有其他一些因素起辅助作用。
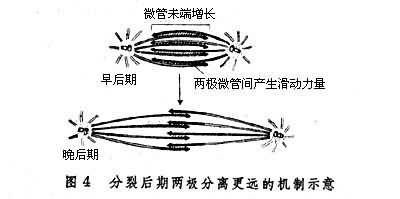
后期染色体运动后期时两组子染色体向两极移动,而在有些细胞两极也被推开更远。关于这种运动的机制尚无定论。后期时着丝点微管在向极的末端不断解聚,因而逐渐变短。这可能是使染色体被拉向两极的重要原因。 因为在体外实验中给模型细胞添加D2O以阻抑微管的解聚时,则染色体向两极移动过程停止,反之,如果添加少量秋水仙素以促使微管解聚速度加快,则染色体向两极移动速度也加快。有些细胞在分裂后期两极分开更远可能是由下述机制造成的:来自两极的极微管在赤道区互相重叠,微管蛋白在它们的自由末端聚合而使微管加长。这些重叠的来自两极的微管互相滑动,使两极推开更远 (图4)。滑动的动力可能是由类动力蛋白 (dynein)ATP酶所产生的。这种分子使ATP水解,释放出滑动所需的能量。这种推测的一个依据是dynein的许多抑制剂,都可阻抑分裂后期的两极分开过程。
与两极的相互离远过程不同,染色体向两极移动过程受dynein抑制剂的影响不显著。推测染色体向两极移动所需能量可能是靠另外一种尚不明了的机制产生的。
持续时间
有丝分裂期所用时间,因生物和细胞的种类不同而有很大差别。一般要1~3小时,但快者有10多分钟就可完成,慢者需5~6个小时,甚至更长时间。通常,胚胎发生早期细胞分裂快,而成体组织细胞分裂慢。
在有丝分裂期中通常是前期和末期时间较长,而中期和后期时间较短。尤其是后期一般需要时间最少。
控制
许多理化因素都影响有丝分裂过程。对于植物和低等动物的细胞,在一定范围内(约8~25℃)温度越低,细胞周期越长,有丝分裂过程也越慢。接近零度的低温可抑制纺锤体的形成,使细胞受阻在有丝分裂中期。电离辐射,如χ射线和γ射线能抑制细胞进入有丝分裂。许多化学药物都有抑制有丝分裂的作用,如秋水仙素、对二氯苯、巯基乙醇、咖啡碱等。促进有丝分裂的化学物质有植物血球凝集素 (PHA)、鲱精胺、激动素、赤霉素等。
在机体内环腺苷酸(cAMP)增加时抑制有丝分裂,而环鸟苷酸(cGMP)增加时促进细胞有丝分裂。
- 参考书目