[拼音]:qunti yichuanxue
[外文]:population genetics
研究群体的遗传结构及其变化规律的遗传学分支学科。它应用数学和统计学方法研究群体中基因频率和基因型频率以及影响这些频率的选择效应和突变作用,还研究迁移和遗传漂变等与遗传结构的关系,由此来探讨进化的机制。
群体指的是孟德尔群体,即一群相互繁育的个体。一个最大的孟德尔群体是一个物种。一个群体中全部个体所共有的全部基因称为基因库。群体中各种基因的频率,以及由不同的交配体制所带来的各种基因型在数量上的分布称为群体的遗传结构。获知了不同世代中遗传结构的演变方式就可探讨生物的进化过程并据以培育各种新的生物品系和品种。可是群体遗传学并不等同于进化遗传学。后者探讨的是物种内变异转化为物种间变异的过程,即物种的形成和绝灭,而前者则仅仅涉及品系间、品种间和亚种间等的变迁。
群体遗传学起源于英国数学家G.H.哈迪和德国医学家W.魏因伯格1908年提出的平衡定律(见遗传平衡)。以后英国数学家R.A.费希尔、遗传学家J.B.S.霍尔丹和美国遗传学家S.赖特等作出了大量的贡献,使群体遗传学发展成为一门独立的学科。
遗传平衡设一对常染色体上的等位基因A和a的频率分别为p和q。假定A对于a为显性,交配是随机的,那么显性个体的基因型及其频率为p2AA+2pqAa,隐性个体的基因型及其频率为q2aa。从而可有三种不同的交配类型(见表)。

从表可见,显性×隐性交配类型中的隐性后代频率
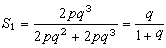
因为q是大于0、小于1的分数,所以S1是1:0和1:1的混合平均数,总是小于1/2。显性×显性交配类型中的隐性后代频率
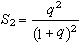
它是1:0和3:1的混合平均数,总是小于1/4。S1和S2称为斯奈德比率。可以看出,在群体遗传学的分析中我们得到的将不是通常孟德尔比率3:1和1:1。而是比它们小的斯奈德比率。
群体遗传学的重要研究指标是考查各种遗传方式使群体达到平衡的速度以及由此所达到的平衡的稳定性。演变只有在这种平衡被打破时才发生;打破平衡的因素有突变、迁移、选择和漂变。
基因突变设μ 是由基因A变为等位基因a的每代突变率(μ=x/n,x是n 代中累积的突变体数)。设群体中A的频率是p。于是下一代A将减少pμ,这样,A的新的频率将是 p(1-μ)。所改变的量 pμ是由突变压造成的,突变压是突变所造成的群体中基因频率改变的度量。如果突变压逐代增加。则基因 A将在群体中逐渐消失,因为
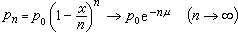
这里 p0是初始频率。如果由a 变为 A的回复突变率是γ,于是基因频率的净变化为△q=pμ-qγ。当这两种相反力量相互抵消时,△q就等于0;得
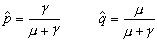
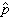
是 A的平衡点,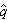 是a的平衡点。平衡的基因频率只决定于相反的两个突变率,而与初始的基因频率无关。由于
是a的平衡点。平衡的基因频率只决定于相反的两个突变率,而与初始的基因频率无关。由于
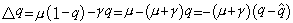 ,
,
每代基因频率的变化与这频率和平衡点的离差有关,当q大于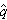 时q就减少,当q小于时q就增加。因此这样的平衡称为稳定平衡,它的变化与交配体制无关。
时q就减少,当q小于时q就增加。因此这样的平衡称为稳定平衡,它的变化与交配体制无关。
一个群体中不同基因型的繁育个体产生数量不等的后代,从而可使群体中基因频率发生变化。设有随机交配群体p2AA:2pqAa:q2aa,其中一部分隐性个体留下的后代数较少,用1-S表示,这里S是选择系数。于是下一代的基因型频率将是p2AA:2pqAa:(1-S)q2aa。而新的隐性基因频率将是[pq+q2(1-S)]/(1-Sq2)=q(1-Sq)/(1-Sq2)。因而每代的q的变化是
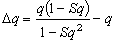
令
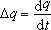
式中t是时间,求积得
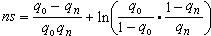
给S 以不同的值,可以算出由已知的q0增加到特定的qn所需要的世代数。通过计算可以看到当q很小时q增加很慢,经过一段时间后增加越来越快,达到顶峰以后又渐趋缓慢。
R.A.费希尔提出了关于自然选择的基本定理。这定理可用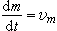 表示,这里m是马尔萨斯氏参数,t是时间(以世代数度量),而υm是参数m的方差。马尔萨斯氏参数所表示的是群体中每个个体的平均后代数,这是适应性的最好度量。它说明在一个群体中只要有大量的变异存在,而且没有其他因素干扰,那么它的适应性就增加。显然这是一种极端的选择论点,把生物界的几乎一切现象都归之于选择的效应。自然选择定理对于单倍体或二倍体,对于随机或非随机交配群体都是适用的。
表示,这里m是马尔萨斯氏参数,t是时间(以世代数度量),而υm是参数m的方差。马尔萨斯氏参数所表示的是群体中每个个体的平均后代数,这是适应性的最好度量。它说明在一个群体中只要有大量的变异存在,而且没有其他因素干扰,那么它的适应性就增加。显然这是一种极端的选择论点,把生物界的几乎一切现象都归之于选择的效应。自然选择定理对于单倍体或二倍体,对于随机或非随机交配群体都是适用的。
繁育个体迁入一个群体或从一个群体迁出都称为迁移。迁移和突变一样会带来基因频率的变化。如果迁入和迁出各是比例m而迁入者中的基因频率是 ,新的基因频率将是q1=q-mq+m=q-m(q-),而△q=q1-q=-m(q-),因而迁移具有和突变相同的效应。如果迁入者的基因频率与本群体相同,则基因频率不起变化。如果一个大群体分成许多小的部分隔离的群体,并且其间没有其他因素的干扰,那么亚群体间的迁移最后将使它们的基因频率都相同,等于整个大群体的基因频率。由迁移造成的基因频率的变化也与交配体制无关。
,新的基因频率将是q1=q-mq+m=q-m(q-),而△q=q1-q=-m(q-),因而迁移具有和突变相同的效应。如果迁入者的基因频率与本群体相同,则基因频率不起变化。如果一个大群体分成许多小的部分隔离的群体,并且其间没有其他因素的干扰,那么亚群体间的迁移最后将使它们的基因频率都相同,等于整个大群体的基因频率。由迁移造成的基因频率的变化也与交配体制无关。
1931年S.赖特提出一个关于群体遗传结构变化的理论,认为当群体是很小时,基因频率在代与代间的随机波动成为群体结构改变的重要因素。假定一个二倍体群体中只有一雌一雄两个个体被保留下来,这时后代的基因型就决定于这一对个体的基因型,不管它们的基因型在选择上是否有利。如果后代因环境改变而大量繁殖,原来被保留的个体就称为建立者。即如果群体很小,选择就不起作用,这时有利的基因也可被淘汰,有害的基因也可被保留。选择不起作用的另一种情况是中性突变,就是既没有利也没有害的突变。日本群体遗传学家木村资生根据许多分子突变(见分子进化)都是中性的这一事实,提出了中性学说,认为中性突变在生物进化中是否被淘汰与选择无关而全凭偶然的机会。这是C.R.达尔文的自然选择学说以外的对生物进化的一种新的解释。
交配体制也影响后代基因型的频率。以随机交配为中点,亲缘关系较近的个体间的交配称为近交或近亲繁殖。近交增加纯合率,血缘越近则纯合率增加得越快,而杂交则正好相反。度量近交程度的一个指标是近交系数,它可以定义为结合的配子之间的相关系数,变异范围由0~1。如以ƒ表示近交系数,则群体中的基因型比率为
AA: p2+ƒpq
Aa: 2pq-2ƒpq
aa: q2+ƒpq如果ƒ保持不变,则群体将达到上式所表达的平衡点。但是如果ƒ逐渐增加,则杂合体逐渐减少,最后将达到0。
由于群体遗传学的理论有它的条件所规定的范围,因而它需要在实验中加以验证,这样便产生了实验群体遗传学。T.多布然斯基的工作是这方面的代表。又由于实验室条件的局限,不能完全反映自然界的情况,就产生了生态遗传学,它把实验室的分析与自然群体的试验结合起来直接验证群体遗传学的理论。英国生态遗传学家E.B.福特的工作是这方面的代表。