[拼音]:youwei mu
[外文]:Caudata, Urodela
两栖纲的1目,终身有尾,幼体及成体体形近似,最不特化。现有8~9科60余属350余种。主要分布于全北界。北美有7科,其中鳗螈科(2属3种、两栖鲵科1属3种)和钝口螈科(4属30余种)为北美特有科;无肺螈科的属种最多(约有27属220余种)在北美洲占优势,中美、南美及南欧只有个别及少数几个属,也是中、南美仅有的科。在古北界,蝾螈科(约有15属50余种)占优势,亚洲东部本科主要分布于东洋界,仅个别属种达东南亚的北缘。小鲵科(共约有8属30余种)为亚洲东部、中亚所特有,最北达北极圈南缘,最南达中国台湾省。隐鳃鲵科(2属3种)分布于亚洲东部和北美东部。非洲热带和大洋洲无有尾类。 中国已知有3科15属和35种左右。小鲵科和蝾螈科的特有属较多。
形态与结构
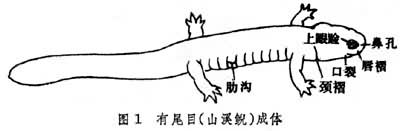
颈部较明显,四肢匀称(图1)(两栖鲵类体长而四肢细弱,鳗螈类只有前肢)。皮肤光滑湿润,紧贴皮下肌肉,全无小鳞。中耳只有耳盖骨-耳柱骨复合结构,从无鼓膜,水栖阶段由耳肌联接肩带,或由韧带联接鳞骨、方骨部位,变态后或陆栖种类靠肩胛提肌的牵引传导声波。一般均不能发声。舌后端不完全游离,不能从后端翻出撮食。上下颌均有小齿,仅鳗螈类覆以角质片。犁骨齿排列形式不一。额骨与顶骨左右各一对,一般没有方轭骨,上颌骨不与方骨相接。椎体双凹型或后凹型,有肋骨。肢带软骨质成分多,肩带仅肩臼周围的部分骨化,左右侧的喙软骨不直接相连,无锁骨;骼骨短,耻软骨与坐骨骈联成平板状,有的有Y形前耻软骨。其形体适于在水中迅速游动,在陆上活动时,躯干很少抬起离开地面,以交替的迈步动作和躯干与尾的波状弯曲而移动前进。能疾走或树栖的种类,其四肢较长,或尾有攀援能力。
生活习性
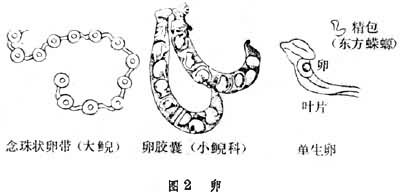
大多生活于淡水水域及其附近。生活在池塘、江河、湖泊、山溪、沼泽中的多为半水栖,其他以终生水栖或陆栖为主。均无交接器;无肛腺者为体外受精,有者为体内受精,其受精方式是:接纳雄性排出的精包,并把精子植入泄殖腔壁的贮精囊内,多为卵生。在水中或水外阴湿处产卵,卵可聚成卵带状、胶囊状或单生(图2)个别种类卵胎生或胎生。幼体上下颌有真齿而不是角质齿;幼体先出前肢芽,有外鳃而无内鳃;平衡枝的有无以及鳃和鳍褶的形状大小,一般与生活习性有关。有若干类群在变态前后,中耳和腭部骨片以及犁骨齿,在形态结构上有颇大的差别。鳃裂封闭是完成变态的主要标志。某些种类或类群的成体保留多种幼态性状,如终生有鳃或有鳃裂,或犁骨齿停滞在幼体期齿列形状等。幼体状态在繁殖方面即已达性成熟阶段者称为童体型或幼态持续型,具永久性幼态性状类群者有隐鳃鲵科、洞螈科、两栖鲵科和鳗螈科,这些终身有鳃或鳃裂的类群,被认为是各自独立发生的;最早的三叠纪童鳃螈亦属之。此外,著名的美西螈在内外因素包括种群于不同分布区的影响下,也可出现童体型。
起源与系统发生
对有尾目的起源主要有两种说法:
(1)直接起源于总鳍鱼类扇骨鱼的孔鳞鱼类(Porole-piformes);
(2)起源于壳椎类, 特别是石炭纪的弛顶螈(Lysorophus),甚至有人把它从细螈目 (Microsauria)中分出, 隶于有尾类最原始的弛顶螈目 (Lysorophia)。这些论点都还没有足够的论据。有尾目最早的化石见于早侏罗世。
关于现生各科间的系统关系:一般把体外受精的小鲵科和隐鳃鲵科(有不少幼态性状),共隶于隐鳃鲵亚目,视之为原始类群。永久性幼态性状而为体内受精者如洞螈科和两栖鲵科,比小鲵科高级,在亲缘系统上与小鲵科较近的是钝口螈科,而钝口螈科又与无肺螈科是近缘。70年代以来,对鳗螈科的分类位置有所争议。有人认为鳗螈类没有肛腺,体外受精,应属原始类群;也有人据其脊神经从最前端椎骨髓弓处穿出,与高级类群蝾螈科相同,认为应属高级类群。如果相似性是平行进化的结果,就成为原始类群之论证;如果体外受精是反向进化的结果(失去体内受精的结构),就成为高级类群的论证。
本目动物摄取动物性食物。再生能力强。食用、药用的种类不多,多用于实验和教学。