[拼音]:zhongqun dongtai
[外文]:population dynamics
一切种群特征的变动(广义的定义),或种群数量在时间和空间上的变动(狭义的定义)。种群动态是种群生态学的核心问题。掌握自然种群动态的规律,便可以更好地计划渔捞量、毛皮兽猎取量,以及野生动物的经济利用等,并对农林业的害虫、害兽以及传播疾病的动物进行有效防治。
研究方法
主要有种群数量统计、实验种群研究和数学模型研究3类。
种群数量统计主要是密度测定,如可测定单位面积和空间上的个体数目。以人口统计为例,可记录每个人的姓名、 住址、出生年月、婚姻和子女状况、 职业、死亡日期和原因等。从这些资料中不仅能获得有关密度和分布的信息,还能得到出生率、死亡率、年龄组成、期望寿命、自然增长率、迁徙等许多重要信息。不过,对野生动物的数量,一般难以进行这么精细的统计。至今只对不太多的动物物种进行过长期、深入的实际调查。另一方面,影响自然种群动态的是综合因素,包括生物的和非生物的以及种群本身内部的因素(见生态因子)。它们共同影响着种群的数量变动,由此造成了对自然种群动态因果分析的复杂性。
实验种群研究即在实验室条件下,控制恒定环境而变动其中一个或几个因素以观察种群生态学特征的变化。例如研究温度、湿度、光照、食物的质和量、以及密度对种群繁殖、存活、行为等的影响。用实验研究方法同样能有效地研究两个种群间的相互作用,如种间竞争、捕食、寄生等。动物实验种群生态学研究,已积累了丰富资料,并在单种种群的增长型及两种混合种群的相互数量关系方面提出了许多很有价值的原理。
实验种群研究的优点是条件控制严格,对结果的因果分析比较可靠,但实验室条件往往与自然种群在野外条件下的生活有区别。动物在实验室条件下可能出现与在自然界中不同的行为,甚至异常行为,如兽类流产率增加、育幼行为改变等。因此,20世纪60年代以来设计了对自然种群实验研究的方法。这类方法兼有自然种群动态调查和实验室研究两种方法的优点。例如在划定的试验区中进行围栏试验,防止种群的扩散;或人工去除自然种群的一部分;或者通过施肥改变种群的食物资源条件等,再观察比较试验区与对照区种群统计参数和种群数量变动情况,从而阐明种群动态的机制。另外,对不同利用强度的种群进行比较研究以获得有关最优种群结构和最优利用方案,这也可认为是一种对自然种群进行实验研究的方法。
数学模型研究利用数学的原理和方法来建立能概括和模拟种群变化的数学公式,并通过实地观察加以验证和修正使之不断完善,借此分析种群变化的内在机制。
设计实验种群研究和数学模型研究,必须以自然种群的动态资料及研究所得的结论为基础,也必须用自然种群的资料检验,以进一步修改和发展模型。实验种群研究,尤其是数学模型研究可协助分析自然种群的动态及其调节机制。
种群数量变动的形式
主要有以下几种:
种群增长有两种基本类型:
(1)J型增长,种群密度按指数迅速增长,遇到环境阻力或其他限制后突然停止增长,通常可用指数方程式描述为dN/dt=rN;
(2)S型增长,开始增长缓慢,经正加速期通过拐点f逐渐放慢(负加速期)而到达某一个极限水平(环境容纳量),通常可用逻辑斯谛方程描述为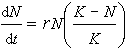 (见种群动态模型)。虽然自然种群增长很少能典型地表现出这两种类型,但可表现为它们的各种变型。
(见种群动态模型)。虽然自然种群增长很少能典型地表现出这两种类型,但可表现为它们的各种变型。
当条件适宜时,某些物种的数量在短期内迅速增加,超过平常的几十甚至几万倍。常见于农林业害虫和害鼠,如蝗虫、粘虫、小家鼠、旅鼠等。蝗虫、旅鼠等在局部地区密度过高时还会发生成群结队大规模迁出,甚至闯入海中。随着种群过剩和环境资源的破坏,大发生后往往继之以种群暴减。一些藻类在水体中也易于大发生,习称“水华”。
生态入侵某些生物因被人类引入适宜地区,种群增大,分布区一步步稳定地扩大。例如欧兔在引进澳大利亚后,每年以70英里的速度扩展分布区,并与牛羊竞争牧草,其数量之大,严重威胁了畜牧业。著名的生态入侵如蟑螂、欧掠鸟入侵北美,中华绒螯蟹入侵欧洲等。另外,致病微生物如流感病毒、鼠疫菌和植物真菌等造成的大流行也都属于生态入侵的范围。
种群下降和灭绝人类在开拓自然时破坏了物种栖息地或持久过度捕猎,都会造成种群数量的逐渐减少甚至种群灭绝。自然种群有时也有持续下降或灭绝的现象。
种群平衡种群数量较长期地维持在一定水平上,没有大起大落,只是围绕一平均密度而上下波动。例如多数稀有种类和大型动物。
季节消长种群数量在一年中随季节的变动称为季节消长。某些小型动物,其短期变动与完成一个世代所需的时间有关;某些海洋动物的种群数量消长可能与潮汐有关;但大部分动物的种群数量波动则是由于环境的季节变化。
不规则波动和周期性振荡自然种群中已知有周期性振荡的有两类:
(1)9~10年周期,主要是美洲兔和猞猁,它们表现出猎物与捕食者的关系;
(2)3~4年周期,最著名的是旅鼠和北极狐,其他还有一些啮齿类、食肉类和鸟类。根据中国兴安岭林区20余年的记录、证实棕背鼠等鼠类具有3~4年周期性,与此周期性相关的还有狐、蜱、红松结实,甚至当地森林脑炎的发病人数。
大多数动物种群波动属于不规则型。不规则波动通常由于非生物因子,尤其是气候因子在不同年份的区别而引起的。曾有人研究为害加拿大橡树的冬尺蠖蛾。通过分析10余年间的生命表,发现该虫的致死因素主要有:
(1)越冬损失;
(2)幼虫被寄生蜂寄生;
(3)幼虫被其他昆虫寄生;
(4)蛹被捕食;
(5)蛹被强姬蜂寄生;
(6)幼虫的微孢子病。根据分析,只有越冬死亡数与总死亡率相平行,这表明它是关键性死亡因素。
种群的空间动态
组成种群的个体在种群内的分布即内分布型大致有 3类:均匀分布、随机分布和聚集分布。它们在自然种群中均有所见,并且已有不少数学模型来描述和检验各种内分布型(见生物分布)。对于许多动物,内分布型研究是正确设计抽样和进行种群统计的基础。
与植物不同,大多数动物是可以运动的。对于在种群分布区内经常移动位置的动物,静态的甚至暂时稳定的内分布型概念是难以适用的。因此,从种群利用空间资源的角度探讨种群的空间关系,可能更具有普遍意义。
任何动物都需要一定的空间来满足其生活的需求。一般说来,种群利用空间资源的方式分两大类:个体(或家庭)的方式和集群的方式。生活于洞穴中的螃蟹、蜥蜴、鼠,在树上单独营巢的鸟,以及狐、鼬等,都是以单体(或其“家庭”)占有一个小区,小区内没有同种的其他个体同时生活。个体(或一个家庭)的领域,有的大,有的小,有的互相联结,有的分开很远,形式多种多样。集群利用空间的方式在动物界中同样很常见:从成群游牧的有蹄类到生活在一起的定居动物群,如旱獭等。每个动物或其家庭都有独立的领域,但有天敌或其他危险物接近时,又通过鸣声或特殊行为彼此报警。
集群使动物能更完善地适应环境。例如,创造有利的小气候(帝企鹅在南极集群进行繁殖),有利于共同捕食(如狼群)、抵御天敌(野牛群、鹿群),合理地利用有限的食物资源(轮流利用领地的游牧群)等。个体(或家庭)的生活方式同样具有适应意义:由于各个体(或家庭)分别占据了各自的领域,减少了种内个体(或家庭)间对食物、隐蔽所等生活条件的竞争,有利于对幼体的抚育和保护,以及度过不利的季节等。
实际上,在自然界中除上述两类利用空间资源的方式外还存在许多中间类型,甚至同一种动物在不同时期内也采用不同方式。例如,许多鸣禽目小鸟在繁殖季节以个体(或家庭)方式生活,而入冬则集合成群,游荡于森林或草地上;一些洄游性鱼类,在进行生殖洄游或索饵洄游时成群游到目的地,然后再以个体(或家庭)形式分散于繁殖基地或索饵场所内。
当动物种群所占有的领域内个体数量过高时,会出现迁移(见迁徙)。物种在其分布区内是以种群的形式存在的,各种群之间保持相对的独立性,但又常通过迁移进行个体互换,这样避免了过分近亲繁殖而使物种退化。
种群调节的机制
种群的数量变动,反映着两组相互矛盾的过程(出生和死亡,迁入和迁出)相互作用的综合结果。因此,影响出生率、死亡率和迁移率的一切因素,都同时影响种群的数量动态。于是,生态学家提出许多不同的假说来解释种群的动态机制。
气候学派气候学派多以昆虫为研究对象,他们的观点主要认为,种群参数受天气条件强烈影响,如以色列学者博登海默认为昆虫的早期死亡率有85~90%是由天气条件不良而引起的。他们强调种群数量的变动,否认稳定性。
生物学派主张捕食、寄生、竞争等生物过程对种群调节起决定作用。例如澳大利亚生物学家A.J.尼科尔森批评气候学派混淆了两个过程:消灭和调节。他举例说明:假设一个昆虫种群每个世代增加 100倍,而气候变化消灭了98%;那么这个种群仍然要每个世代增加一倍。但如果存在一种昆虫的寄生虫,其作用随昆虫密度的变化而消灭了另外 1%,这样种群数量便有了调节并能保持稳定。在这种情况下,寄生虫消灭的虽少却是种群的调节因子。由此他认为只有密度制约因子(见生态因子)才能调节种群的密度。
50年代气候学派和生物学派发生激烈论战,但也有的学者提出折衷的观点。如A.米尔恩既承认密度制约因子对种群密度的决定作用,也承认非密度制约因子具有决定作用。他把种群数量动态分为3个区,极高数量、普通数量和极低数量。在对物种最有利的典型环境中,种群数量最高,密度制约因子决定种群的数量;在环境条件对物种很不利、变动很激烈的条件下,非密度制约因子左右种群数量变动。这派人认为,气候学派和生物学派的争论,反映了他们工作地区环境条件的不同。
一般说,种群调节机制同生态策略有关;栖居于不稳定环境中的r 策略者更多地受非密度制约因子的作用,而栖息于有利的稳定环境中的 K策略者则更多地经受密度制约因子的调节。
食物因素强调食物因素的学者也可归入生物学派。如英国鸟类学家D.拉克认为,就大多数脊椎动物而言,食物短缺是最重要的限制因子。自然种群中支持这个观点的例子还有松鼠和交嘴鸟的数量与球果产量的关系、猛禽与一些啮齿类数目的关系等。
强调食物因素为决定性的还有F.A.皮特尔卡的营养物恢复假说。这一假说说明冻原旅鼠的周期性变动:在旅鼠数量很高的年份,食物资源被大量消耗,植被量减少,食物的质(特别是含磷量)和量下降,幼鼠因营养条件恶化而大量死亡以致种群数量下降。低种群密度使植被的质和量逐步恢复,种群数量也再度回升,周期大约为3~4年。
自动调节学说上述各种学说都集中在外源性因子,主张自动调节的学者则将研究焦点放在动物种群内部,其特点包括:强调种内成员的异质性,异质性可能表现在行为上、生理特征上或遗传性质上;认为种群密度的变化影响了种内成员,使出生率、死亡率等种群参数变化;主张把种群调节看成是物种的一种适应性反应,它经受自然选择,带来进化上的利益。按所强调的特点,又分为行为调节、内分泌调节和遗传调节等学说。
(1)行为调节:英国的V.C.温-爱德华兹认为动物社群行为是调节种群的一种机制。以社群等级和领域性为例。社群等级使社群中一些个体支配另一些个体,这种等级往往通过格斗、吓唬、威胁而固定下来;领域性则是动物个体(或家庭)通过划分地盘而把种群占有的空间及其中的资源分配给各个成员。两者都使种内个体间消耗能量的格斗减到最小。使空间、资源、繁殖场所在种群内得到最有利于物种整体的分配,并限制了环境中的动物数量,使食物资源不致于消耗殆尽。当种群密度超过这个限度时,种群中就有一部分“游荡的后备军”或“剩余部分”,它们一般不能进行繁殖,或者被具有领域者所阻碍,或者缺乏营巢繁殖场所。这部分个体由于缺乏保护条件和优良食物资源也最易受捕食者、疾病和不良天气条件所侵害,死亡率较高。这样,种内划分社群等级和领域,限制了种群不利因素的过度增长,并且这种“反馈作用”随着种群密度本身的升降而改变其调节作用的强弱。
(2)内分泌调节:J.J.克里斯琴最初用此解释哺乳动物的周期性数量变动,后来这理论扩展为一般性学说。他认为,当种群数量上升时,种内个体经受的社群压力增加,加强了对中枢神经系统的刺激,影响了脑下垂体和肾上腺的功能,使促生殖激素分泌减少和促肾上腺皮质激素增加。生长激素的减少使生长和代谢发生障碍,有的个体可能因低血糖休克而直接死亡,多数个体对疾病和外界不利环境的抵抗能力可能降低。另外,肾上腺皮质的增生和皮质素分泌的增进,同样会使机体抵抗力减弱,而且由于相应的性激素分泌减少,生殖将受到抑制,致使出生率降低,子宫内胚胎死亡率增加,育幼情况不佳,幼体抵抗力降低。这样,种群增长因上述生理反馈机制而得到抑制或停止,从而又降低了社群压力。
(3)遗传调节:英国遗传学家E.B.福特认为,当种群密度增高时,自然选择压力松弛下来,结果是种群内变异性增加,许多遗传型较差的个体存活下来;当条件回到正常的时候,这些低质的个体由于自然选择压力增加而被淘汰,于是降低了种群内部的变异性。他指出,种群的增加必然为种群的减少铺平道路。
D.奇蒂提出一种解释种群数量变动的遗传调节模式。他认为,种群中的遗传双态现象或遗传多态现象有调节种群的意义。例如在啮齿类中有一组基因型是高进攻性的,繁殖力较强,而另一组基因型是繁殖力较低,较适应于密集条件的。当种群数量初上升时,自然选择有利于第一组,第一组逐步替代第二组,种群数量加速上升。当种群数量达到高峰时,由于社群压力增加,相互干涉增加,自然选择不利于高繁殖力的,而有利于适应于密集的基因型,于是种群数量又趋下降。这样,种群就可进行自我调节。可见,D.奇蒂的学说是建立在种群内行为以及生理和遗传的变化之上的。
组件生物的种群过程
植物种群生物学是70年代以来新发展的一个领域;它把生物划分为两大类:单型生物和组件生物。传统的种群生态学主要讨论单型生物的种群动态。单型生物的种群大小可以用个体数量来描述,它们都是由单一合子发展起来的,但对于树和草等高等植物的种群,只用植株数很难描述种群的动态过程。有些树株发育着茂盛的枝叶,有些则株条稀疏;有的草生长成片,草根互相交错联结,甚至难以分清哪一些属于哪一个个体以及是由哪一个合子发育而成的。它们虽然与单型生物不同,但却有一个共同的特点,即其组成单位重复地出现。通常由一个合子发展起来的一系列重复出现的组成单位可以称为组件,因此,这类生物叫作组件生物。
单型生物的种群可用个体数量“N”来描述;而组件生物的种群通常有两个明显不同的层次:一是遗传个体的数量,它与单型生物的个体相当;二是每一遗传个体上组件的数量,它是单型生物所没有的。通常可用“N”表示遗传个体的数量,“η”表示组件的数量。例如,在池中培养浮萍,如果培养由一个合子开始,经过一段时间的增长以后,池中就有了从一个遗传个体分生出来的浮萍叶状体(组体)组成的种群。这个 η水平的种群增长是由新组件产生和旧组件死亡的过程所决定的。如果培养是由若干个合子开始的,那么池中就包括有η和N两个水平的种群,η=N×平均每个个体含有的组件数。
组件生物种群的年龄结构,除N水平的以外,对组件生物的组件还有 η水平的。一株草、一棵树所生的叶和芽是不同时间产生的。
组件生物的种群结构具有两个水平,同样也有两个水平的多态现象。传统的多态现象代表了N 水平的遗传个体之间的差别,例如雌雄异株植物的性多态现象及同一种植物的颜色多态等。η水平的多态现象,如薮枝虫营养螅体和生殖螅体的分工,树上的长枝与短枝,以及雌雄同株植物上的雌花与雄花等。
组件生物的组件构筑型,即重复出现的组件在空间上的配置型式,在决定它们的适应能力上具有重要意义。单型生物的许多重要特性,诸如摄食、求偶、竞争、逃避捕食动物等,往往与它们的运动及其他行为有关。但是组件生物是营固着生活的,光、水、矿质营养物等资源多从根、茎、叶的直接周围空间获取,上述这些生命功能的完成与组件的构筑型有密切关系。例如花的位置应容易被传粉昆虫发现,枝系的配置应利于最有效地搜集光辐射,根系进入土壤的区域应是水分最多的等,这些特征都包含着生长的方向和组件的布局问题J.L.哈珀认为,植物或其他组件生物的理论生态学的进一步发展,应该把注意力更多地放在遗传个体和组件的空间位置上,而不是放在更加一般的密度关系方面。