[拼音]:danao
[外文]:cerebrum
又称端脑,脊椎动物脑的高级的主要部分,由左右两半球组成,在人类为脑的最大部分,是控制运动、产生感觉及实现高级脑功能的高级神经中枢。脊椎动物的端脑在胚胎时是神经管头端薄壁的膨起部分,以后发展成大脑两半球,主要包括大脑皮层和基底核两部。大脑皮层是被覆在端脑表面的灰质、主要由神经元的胞体构成。皮层的深部由神经纤维形成的髓质或白质构成。髓质中又有灰质团块即基底核,纹状体是其中的主要部分。广义的大脑指小脑幕以上的全部脑结构,即端脑、间脑和部分中脑(见中枢神经系统)。
脊椎动物的端脑,不论在外形或内部构造方面,在种系发展中均有很大的变化(图1、图2)。圆口纲动物端脑包括 3部分:
(1)嗅球及膈,只接受嗅觉传入神经。
(2)纹状体,其位置一般在腹外侧部或背外侧部。
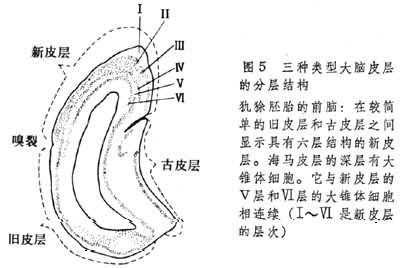
(3)大脑皮层,它的位置一般在背外侧部,此部在动物以后的发展过程中覆盖在其他神经组织的外表,故称大脑皮层(图1)。两栖动物的大脑皮层可分 3部。内层为古皮层或海马的原基。外层的是旧皮层或梨状区。在两层之间有一个背侧区。一般认为在动物发展过程中此区即相当背侧皮层或新脑皮层,相当于哺乳动物的新皮层(图3)
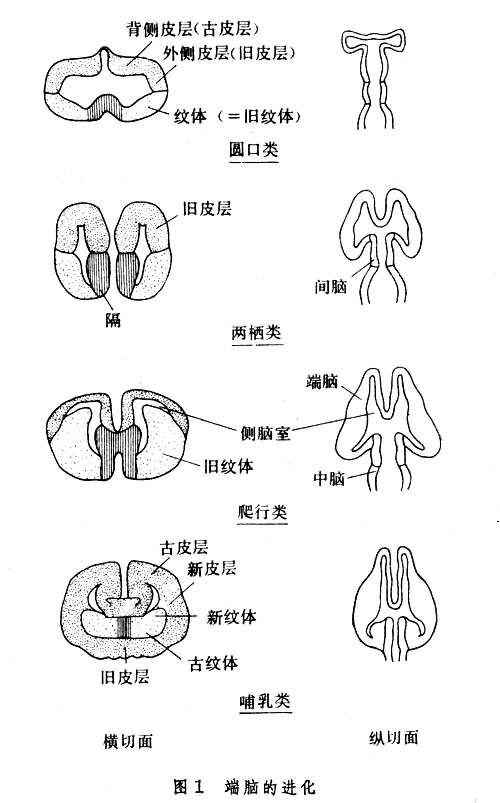
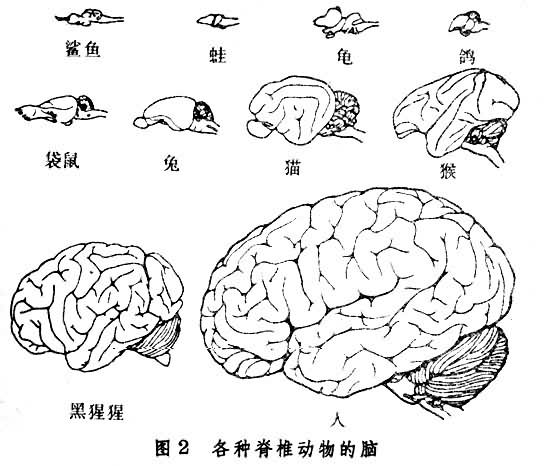
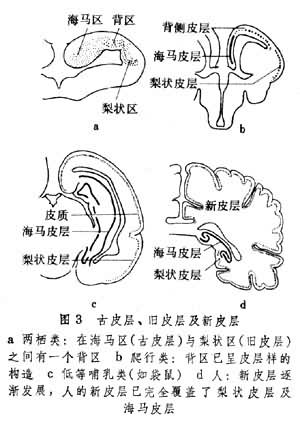
在低等脊椎动物,大脑皮层的灰质是贴近脑室的,大脑的表面由神经纤维(白质)覆盖。直到两栖动物都保持着这种格局。从爬行动物开始,端脑某些部分的神经细胞开始向浅层转移,形成清晰的表浅灰质,即原始皮层。由于上行、下行投射纤维的不断增多,更使灰质向表浅部迁移。哺乳动物的整个大脑表面都变成特化的、分层的灰质。也有一些灰质块埋藏在大脑白质之中,这就是基底核(基底神经节)。大脑的发展充分地说明了动物进化中神经系统的(功能)大脑化过程。动物越高等, 对大脑的依赖性就越大。切除大脑皮层的鼠,它的活动行为粗看起来还大体正常;而一个无皮层的人则成为完全的残废。大脑皮层在皮层下神经核及传入、传出系统的协同下,接受和分析感觉信息,设计并控制运动,调节着内环境恒定过程,储存记忆的经验。对一些复杂的脑功能,如思考、计算、判断等,大脑皮层是最重要的神经结构。但是,即使这些复杂功能,大脑皮层也必须得到其他部分的配合。所以正常的行为是完整的神经系统功能的总的体现。
哺乳动物啮齿类的大脑表面平坦,无明显沟回。狗、猫等较高等食肉目动物的大脑皮层,由于褶皱而出现许多沟回,因而大大地增加了皮层的表面积。人类大脑皮层表面积一般可达3200~2500平方厘米,厚约1.5~4.5毫米,体积约600立方厘米,其中含12~140亿神经元和更多的神经胶质细胞。大脑皮层是一个分层的神经组织。哺乳动物的大部皮层由表及里可分6层:
(1)分子层;
(2)外粒层;
(3)外锥体层;
(4)内粒层;
(5)内锥体层(神经节细胞层);
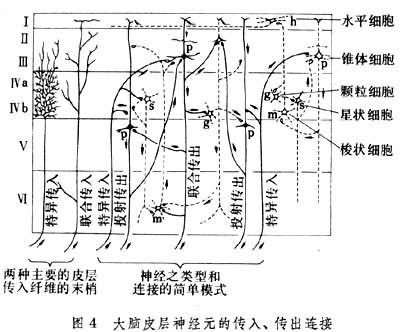
(6)多形层。在这6层中主要分布着3种类型的神经细胞,即锥体细胞、星形细胞和梭形细胞,在中央前回第4区第5层有巨大的锥体细胞,叫做贝茨氏细胞。星形细胞主要司大脑皮层内的连接。投射纤维、联络纤维和连合纤维主要由锥体细胞发出(图4)。由大脑皮层走向其他脑区的纤维叫做投射纤维,分别终止于脊髓、脑干的运动神经核、桥核、尾状核、壳核、脑干网状结构;少数纤维可终止于脊索核、下橄榄核、四叠体等处。另外还有很多皮层-丘脑纤维,其终止区常是发出相应丘脑皮层纤维的所在地。在同侧大脑皮层不同脑区之间相互联系的纤维叫做联络纤维。连接两侧大脑半球的纤维叫做连合纤维。大脑皮层的传入纤维主要发源于丘脑。近来发现由蓝斑发出的儿茶酚胺能纤维和由中缝核群发出的5-羟色胺能纤维部分终止在皮层细胞。由于皮层的传入、传出纤维关系非常复杂,加上神经细胞的类型又多,所以他们之间的突触连接很难分辨。乙酰胆碱、去甲肾上腺素等都是它的递质;此外γ-氨基丁酸被认为是大脑皮层特异的抑制性递质。70年代以来发现一些活性脑肠肽如血管活性肠肽(VIP)、胆囊收缩素(CCK)等也可能是大脑皮层的重要递质。
大脑皮层有古皮层,旧皮层,新皮层之分。古、旧皮层的分层不清楚,所以叫做不均皮层、未层次化皮层、异质皮层、异生皮层,而新皮层是6层型的叫做同形皮层或均质皮层、同生皮层(图5)。
大脑皮层的分区
各种细胞层的厚度、细胞的排列、型式等叫做细胞构筑或细胞结构;纤维、血管、化学构成等的特点分别叫做纤维构筑、血管构筑和化学构筑。
不同作者根据大脑皮层各区的细胞构筑不同,把大脑皮层分为若干区,K.布罗德曼把人的大脑皮层分成47个区叫做布罗德曼氏皮层区。这种分区虽然带有一些人为的性质,但它一方面给出了脑区的具体编号,便于研究和描述;另一方面细胞构筑的不同也多少反映皮层功能的特点,如17区与视觉关系密切,也叫视觉皮层,4区与身体各部的运动功能有密切关系所以也叫运动区皮层等。
大脑皮层的感觉、运动功能
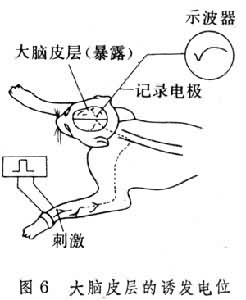
大脑皮层具有控制运动和分析感觉信号的功能。这方面的知识大都是通过刺激或损毁实验和临床观察等方法获得的。如用电刺激动物皮层的一定部位,观察动物的反应,从而了解这一部位的功能。或者用物理和化学的方法破坏皮层某一局部组织,观察是否引起某一特定功能的消失。在感觉投射的研究中,特别是大脑皮层感觉区的定位中,常用诱发电位的方法。所谓诱发电位一般是指感觉系统受到刺激时,在中枢神经系统内引起的电变化。大脑皮层的诱发电位,则是指感觉传入系统受到刺激时,在皮层上某一局部区域所引出的电变化 (图6)。用微电极技术,可以记录单个神经细胞的活动。利用抗原、抗体特异反应的原理发展起来的放射免疫、免疫组织化学、免疫酶标等技术,为脑中化学物质、神经连接等方面的研究开辟了新的途径。
 感觉皮层
感觉皮层
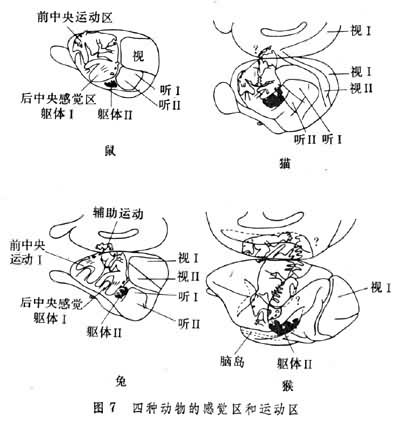
大脑的感觉皮层是指接受丘脑特异转换核传入的大脑皮层区域(图7),如视区、听区、体感区等。内脏感觉区的丘脑传入反射不如上述几区那样清楚,一般不叫做内脏感觉区皮层。
(1)体表感觉区皮层(中央后回第3,1及2区)。全身体表感觉的主要投射区。通过灵长类动物皮层诱发电位的研究,发现向中央后回的感觉投射有以下特点;体表感觉传向皮层的投射是交叉的,即一侧的体表感觉传到对侧皮层的相应区,但头面部感觉的投射是双侧性的;投射区有一定的空间分布,下肢代表区在顶部,上肢代表区在中间部,头面部代表区在底部,因此,总的排列是倒置的;投射区的大小与不同体表部位感觉灵敏程度有关,例如拇指和食指的代表面积要比胸部十二根脊神经传入的代表区总面积大几倍,这种结构特点有利于精细的感觉分析。脑外科手术时,用适宜强度的电流刺激病人大脑皮层,观察到刺激中央后回顶部可以引起似乎来自下肢刺激的主观感觉,刺激中央后回底部可以引起似乎来自面部的主观感觉。
除中央后回第1感觉区外,在猫、猴等动物以及人还发现有第二感觉区。人的第 2感觉区位于中央前后回的最下部。体表感觉在第 2感觉区的投射也有一定空间分布,但这种分布属于正立的而不是倒置的。在人脑,刺激第2感觉区可引起体表一定部位的麻木感,这种感觉具有双侧性。从种系发生上看,第2感觉区似较原始,只能对感觉作粗糙的分析。人在切除第二感觉区后,并不产生显著的感觉障碍。
(2)肌肉本体感觉区。中央前回是运动区,也是肌肉本体感觉投射的代表区。切除动物的运动区,由本体感觉刺激作为条件刺激所建立的条件反射就发生障碍。刺激人体中央前回,也会引起受试者企图发动肢体运动的那种主观感觉。
(3)视觉区皮层。枕叶皮层是视觉的投射区,左侧枕叶皮层接受左眼颞侧视网膜和右眼鼻侧视网膜的传入纤维投射,右侧枕叶皮层接受右眼颞侧和左眼鼻侧视网膜的传入纤维投射。枕叶皮层的视觉代表区和具体位置在皮层内侧面的距状裂(距状沟)上、下缘。视网膜上半部投射到距状裂上缘,下半部投射到下缘;视网膜中央凹的黄斑区投射到距状裂前部。电刺激人脑距状裂上、下缘,可以使受试者产生简单的主观光感觉,但不能引起完整的视觉形象。
(4)听觉区皮层。颞叶皮层是听觉的投射区域。听觉的投射是双侧性的,即一侧皮层代表区与双侧耳蜗感受器的传入有关。猫的听觉皮层代表区位于外雪氏回,耳蜗一定部位的感觉传入投射到听觉代表区的相应点。人的听觉皮层代表区位于颞横回和颞上回(第41、42区),电刺激上述区域,能引起受试者产生铃声样或风吹样的主观音感觉,在第41区还有不同音频感觉投射的分野;高音感觉区在此区的后内侧,低音感受区在此区的前外侧。
运动区皮层大脑皮层与运动有关的区域有:中央前回运动区、辅助运动区、第2运动区(图7)。中央前回运动区叫做运动区皮层,包括布劳德曼氏皮层区第 4区和第6区。第4区主要与远端关节如手指、脚趾等精巧的动作,乃至个别肌肉的动作有关。第 6区主要与近端关节如肩、髋等关节运动有关。运动区皮层有下列功能特点:
(1)对躯体运动的管理是对侧性的,即一侧运动区皮层控制另一侧躯体的肌肉运动,但头面部肌肉的运动,如咀嚼、喉及脸上部运动的支配都是双侧性的。
(2)具有精细的功能定位,即皮层的一定区域支配身体一定部位的肌肉呈倒立的“小人”样分布。这种情况叫做躯体定位。
(3)身体不同部位在皮层的代表区,其大小与肌肉的大小无关,而与肌肉运动的精细和复杂程度有关。如拇指所占的皮层面积差不多是大腿所占皮层面积的10倍。运动区皮层最重要的功能是管控制随意运动。某些复杂的运动反射必须有运动区皮层参加。另外,在中央前回与岛叶之间有第二运动区,在大脑皮层额上回沟内侧面(两半球纵裂的侧面)有辅助运动区。目前,这两区的生理功能还不很清楚,刺激一侧辅助运动区及第二运动区,能引起两侧躯体运动反应,但需用较强电刺激。
大脑皮层功能的柱状结构
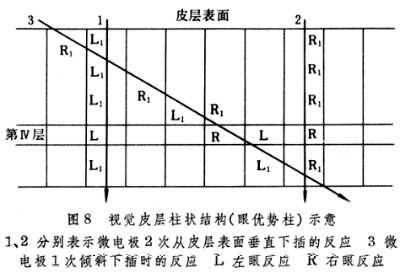
V.B芒卡斯尔认为大脑不但在分区上,而且在皮层深浅各层的功能上也有一定的排列和连接方式。皮层细胞在深浅方面上的柱状排列构成皮层的最基本功能单位,垂直于脑表面的直径约1毫米左右的细胞柱具有相同的功能。皮层中神经元的联系主要是垂直向的联系,这些神经元链插入在输入和输出之间,形成复杂的环路进行信息处理。如经丘脑特异核的传入冲动经第Ⅳ层的颗粒细胞,向下到第Ⅴ、Ⅵ层,向上到更表浅的第Ⅱ、Ⅲ层细胞,经垂直向神经元环路的整合后输出。每一细胞柱,经回返抑制的作用还可压抑邻近细胞柱的活动,从而进一步突出垂直部分的神经活动。已知运动皮层、视觉区皮层和听觉区皮层都有这种柱状组构特点(图8)。柱状组构的结构基础目前还不十分清楚。但近年发现,小鼠口旁的20多根刚毛,在皮层体感区上每根均有相应的成桶状的神经细胞群。
大脑皮层的联络区
联络区是指那些在刺激、损毁及电生理实验中未能发现明显的运动、感觉或自主性功能变化的那些大脑皮层区。这些区在高等动物更为发达,灵长类的皮层联络区主要指位于顶叶、颞叶、枕叶之间的顶-枕前区皮层以及位于运动区之前的前额叶皮层。从神经联系上看,这些脑区不接受丘脑中继核的投射,而只接受那些丘脑联络核的传入投射。丘脑联络核由于主要接受丘脑内部相互的连接,而不接受丘脑以外来的传入投射,因而得名。所以这些相应的大脑皮层也就被命名为联络区皮层,也叫“内源性皮层”。
对人类及高级灵长类动物联络区的损伤和刺激实验说明,联络区的功能一方面与其邻近的初级感觉区皮层功能有较密切的关系,如顶-枕叶皮层与体感的知觉及行动关系较密切,颞中区与听觉的关系较密切,下颞区与视觉功能较密切。另一方面联络区的功能已经超出了特异感觉的范围。例如顶-枕联络区的作用似乎是将事物的空间排列与机体本身的形状和位置加以比较,根据身体的内部状态所处的条件来确定外界事物的意义,顶叶损伤病人的症状有半边身体不复存在的感觉。有的可能与短期记忆功能有关。
大脑皮层与条件反射
见条件反射。
大脑半球与学习、记忆
见学习和记忆。
大脑皮层功能两侧不对称性和人类的语言功能
人大脑皮层的一定部位受伤时,会出现语言障碍。损坏左侧额下回的布罗卡氏区 (第44区)可出现运动失语症。这种病人有关发音的肌肉运动并无障碍,但不会说话。损坏左侧颞上区的韦尼克氏区(第22区)时,会出现感觉失语症。W.G.彭菲尔德等在给病人作手术时刺激其大脑 皮层,发现也正是这些区的刺激可以使清醒病人的言语停顿或发生障碍,证实了过去临床的观察结果(图9)。
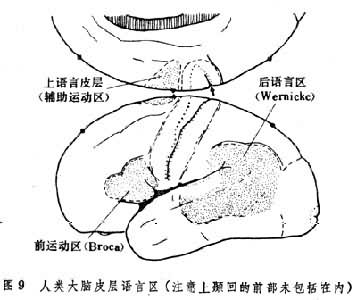
左侧或右侧大脑皮层受伤对于语言功能的影响是完全不同的。J.N.杰克逊1869年最先提出“领导半球”的看法。他注意到一个惯用右手(右利)的人,如右侧偏瘫,会出现失语症;左侧偏瘫则不出现。于是就有人把右利者的左侧大脑半球叫做优势半球或主要半球,而其右侧半球为次要半球。以后发现,哪个半球与语言有关,完全是遗传因素所决定的。至于用手的右利或左利除遗传因素外,还有后天因素。一个左利者发生右侧偏瘫时,同样会产生失语症。
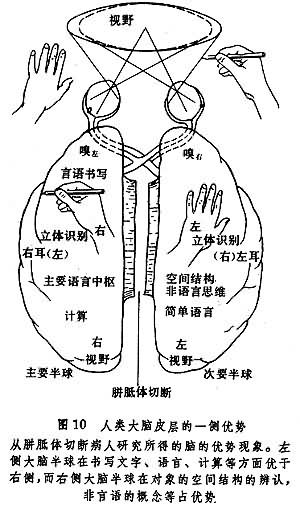
所谓优势,并非一切功能都是在一侧占优势。R.W.斯佩利等在切断胼胝体的“裂脑”人身上进行了细致的观察,发现多数人的左侧大脑半球在语言、抽象思维等方面占优势,而右侧大脑半球在非语言功能、空间辨认等方面占优势。说明大脑两半球的功能不完全对称而有所分化(图10)。
基底神经节
鱼类及两栖类动物的端脑的腹外侧壁是纹状体。到了爬行动物才分成古纹状体和旧纹状体。哺乳动物的杏仁、苍白球属于古、旧纹状体,而尾核、壳核则属于新纹状体,这些结构的全部统称基底核(基底神经节)。底丘脑核、黑质、红核等在功能上与纹状体有密切关系。
大脑皮层不发达动物的基底神经节是控制运动的主要中枢。在皮层高度发达以后,基底神经节的功能主要与运动调节有关,是锥体外系的主要组成部分。人基底神经节的病变,常伴有运动障碍,如震颤麻痹的损害在纹状体或黑质,其主要症状是面部表情木呆、步态死板,肌肉僵直等。基底神经节本身并不直接发出下传通路,它对运动的调节与下述环路有关(图11)。

已知多巴胺能神经元胞体在黑质,而尾状核、壳核是黑质、纹状体投射的末梢所在。震颤麻痹病人的黑质及或尾状核、壳核的多巴胺含量明显降低,是由于损害了上述投射通路。临床上应用左旋多巴,可以补偿这类病人多巴胺的降低,故对震颤麻痹病有效。纹状体除含多巴胺外,还含乙酰胆碱、5-羟色胺、γ-氨基丁酸等递质,这些递质含量的增减,与震颤麻痹的发病和缓和有一定关系。
杏仁核属于古纹状体,它的功能主要与情绪及内脏活动的调节有关(见边缘系统)。
- 参考书目
- 陈宜张:《神经系统电生理学》,人民卫生出版社,北京,1983。
- T.H.Bullock et al., Introduction to Nervous Systems,Freeman & Co.,San Francisco, 1977.