[拼音]:xibaomo shouti
[外文]:cell membrane receptor
细胞表面的一种或一类分子,它们能识别、结合专一的生物活性物质(称配体),生成的复合物能激活和启动一系列物理化学变化,从而导致该物质的最终生物效应。
“受体”这个名词在19世纪末20世纪初已经开始应用,当时认为药物作用时必须与靶细胞上与之互补的分子──受体──相结合后才能发挥效应。后来受体概念的应用逐渐扩展到药物以外的其他信息分子作用于细胞的过程。
化学组成和结构
迄今所知,受体几乎都是蛋白质(糖蛋白,脂蛋白或糖脂蛋白)。
由于细胞内受体含量极微,有些受体稳定性又差,因此受体的分离、纯化比较困难。迄今只有从电鳐和电鳗的电器官中分离的乙酰胆碱的烟碱胆碱能受体和从正常人胎盘中分离的胰岛素受体已经得到纯度很高、数量足够的样品,因而对它们的结构也有了较多的了解。电鳐的乙酰胆碱的烟碱胆碱能受体是由分子量为26000~64000的四种亚基组成,从电鳗分离的受体则由分子量为42000和54000二种亚基组成。电子显微镜观察的结果表明,这种受体由5~6个直径为3~4纳米的亚基构成玫瑰花结状结构,其直径约8~9纳米,中心可能是离子通道。胰岛素受体的斯托克半径约68~72埃,分子量约30~100万,它主要由3种亚基:α(分子量约135000),β(分子量约95000)和γ(分子量约45000)组成。α亚基为结合亚基,亚基之间靠-S-S-相连,亚基内部还有-S-S-结构。
数量和分布
一种细胞膜可以含有几种不同的受体,如脂肪细胞膜上含有肾上腺素、胰高血糖素、胰岛素等近10种激素受体。它们的数目互不相同。同一受体在不同细胞膜上的受体数目也是不同的。一般的受体的密度为103~104个/细胞,但电鳐电器官上的乙酰胆碱的受体的密度和数量较大,分别为104~105微米2或1011个/细胞。受体的数目在正常生理条件下是恒定的,但由于细胞生理状态不同(如生长速度,分化程度,细胞周期等)和外界环境变化的影响,也会发生一定的改变。
受体在膜上的分布是不均匀的,经常相互聚集形成“簇”的形式。
调节
受体调节的因素和途径很复杂,在正常生理情况下受体数目受微环境影响而上升或下降,称为上升或下降调节。其中与受体结合的配体浓度对调节受体具有较重要的作用,例如,当动物或人的血液中胰岛素浓度较高时,靶细胞上的胰岛素受体浓度即下降,如果胰岛素浓度降低时,受体数目会迅速上升。受体的调节还可通过“负协同效应”来进行,即:受体-受体的相互作用会导致受体的亲和力发生变化。例如,受体与胰岛素分子的结合会促使受体与周围其他受体之间的相互作用,导致受体的亲和力下降,从而使胰岛素分子从受体上解离下来。
抗体
在机体内已经发现某些受体的自身抗体,例如,1975年美国从一种β型严重胰岛素抵抗症病人中发现有胰岛素受体的自身抗体。这些抗体与受体结合可模拟胰岛素的许多作用(例如,抑制脂肪分解,刺激葡萄糖的转移和利用),但它会逐渐降低细胞对受体被结合后的生物化学反应的敏感性。加之抗体的存在也会降低受体对胰岛素的亲和力,结果在临床上就表现为严重的胰岛素抵抗症。
除胰岛素受体外,儿茶酚胺受体,乙酰胆碱的烟碱胆碱能受体,促甲状腺素受体在病理情况下都发现有相应的自身抗体存在。
传递信息的机理
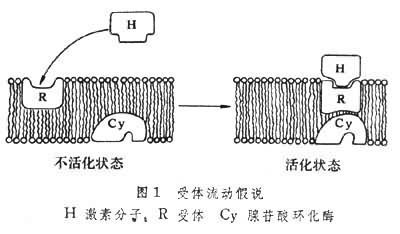
激素受体激素与受体结合后如何产生生物效应?60年代提出的第二信使假设认为,作为第一信使的激素分子与细胞膜受体结合后并不进入细胞。结合激素的受体能使位于膜上的腺苷酸环化酶活化,从而使ATP转成环(化)腺苷酸(cAMP),后者称为第二信使,它能引发细胞内一系列生化反应而产生最终生物效应。例如,肾上腺素在肾上腺髓质分泌后通过血液输送至肝细胞产生效应。它们与肝细胞表面受体相结合后能使膜上腺苷酸环化酶活化催化ATP形成cAMP,后者使蛋白激酶等一系列酶蛋白相继活化,最终使糖原分解成葡萄糖,从而使血糖浓度升高。其他一些激素,例如,促肾上腺皮质激素(ACTH),促卵泡激素(FSH),胰高血糖素和儿茶酚胺激素等在作用时也都是分别与它们的靶细胞受体相结合,导致腺苷酸环化酶的激活产生cAMP。这些激素在细胞膜上具有各自专一的受体,但腺苷酸环化酶却是共用的。待激素分子与膜受体相结合形成复合体后,它们在膜上进行侧向移动,一旦与腺苷酸环化酶相遇偶联才能导致后者的构象变化而被激活。一旦激素稀释或解离,受体和腺苷酸环化酶便又经移动而回到非偶联状态。这称为“受体流动假说”(图1)。
有些受体,如α类肾上腺素能受体与激素结合后细胞内不是cAMP而是Ca2+的浓度有明显增加,这可能是由于膜上Ca2+通道开启,使细胞外Ca2+进入细胞引起的。Ca2+能与钙调蛋白结合并引起一系列反应,因而有人认为Ca2+也是一种第二信使。
近年来,发现有些激素,如胰岛素,与细胞膜受体结合后并不仅仅停留在表面,形成的胰岛素-受体复合物可通过内吞方式进入细胞内部(见内吞与外排),继而这些复合物再与溶酶体融合或与其他细胞器膜相结合而产生生物效应。
利用高纯度的胰岛素研究表明,胰岛素与受体的α亚基结合后可使受体本身固有的某种蛋白激酶活化,使受体分子本身,主要是β亚基发生磷酸化作用,这称为自身磷酸化作用,它继而又引起一连串的磷酸化反应,这可能是胰岛素产生生物效应过程中一个十分重要的环节。
神经递质受体神经递质有十多种,它们各自有一种或一种以上的受体。就乙酰胆碱而言,在脊椎动物中至少有三种受体,其中烟碱胆碱能受体和蕈毒胆碱能受体研究得比较多。烟碱胆碱能受体分布于自主神经节、中枢、电鳗的电器官等的细胞膜中,当受体与烟碱结合而被激活后,离子通道很快开启,开启的持续时间短(毫秒级)。蕈毒胆碱能受体存在于副交感神经、平滑肌等组织中,当受体与蕈毒结合而被激活时,离子通道开启缓慢,持续时间较长(秒级)。上述二种受体由于各具有与乙酰胆碱不同的基团相匹配的结构因而都能与之相结合。
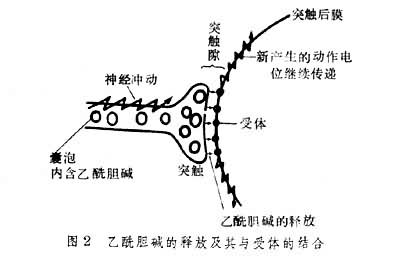
在中枢神经系统,前一神经原的轴突末梢兴奋时,突触处细胞质内大量储存泡通过与突触前膜相融合而将所含的乙酰胆碱释放至距离为0.5纳米的突触隙,之后乙酰胆碱与突触后膜上的受体相结合,使突触后的神经原发生兴奋,从而使神经冲动传递下去(图2)。释放的乙酰胆碱如果作用于效应器(如肌肉),就能引起反应(如收缩)。当乙酰胆碱与肌肉细胞受体结合后,会引起它的构象发生变化,打开离子通道,使膜对Na+、K+的通透性瞬时增加,从而导致膜电位的去极化,使肌肉收缩。一个与乙酰胆碱结合了的受体可导致5×104离子通过,显示了受体的放大作用。
 毒素受体
毒素受体
现在发现很多毒素也是通过与细胞膜上的受体相结合后才产生效应的。如霍乱毒素是霍乱弧菌产生的外毒素,分子量为84000,由A、B二种亚单位组成。A亚单位有两条肽链A1和A2,由一对二硫键联接。亚单位B与细胞膜上的受体相结合。亚单位A1则具有激活膜上腺苷酸环化酶的作用。
霍乱毒素的受体是一种神经节苷脂,毒素与它结合后可能会发生一系列反应,如首先引发受体的构象变化,接着亚单位A1在激活腺苷酸环化酶的过程中将烟酰胺腺嘌呤二核苷酸 (NAD)中的腺二磷核糖转移到细胞膜中的一种蛋白质上。在正常情况下,鸟三磷通过与这种蛋白质相结合使腺苷酸环化酶激活,待鸟三磷被鸟三磷酶水解,激活作用即停止。但是如果鸟三磷结合在含有腺二磷核糖的蛋白质上就不易被水解,因而延长了腺苷酸环化酶作用的时间,产生的生物效应的持续时间也比较长。
霍乱毒素的作用与激素有几点不同:
(1)激素的启动时间很短,霍乱毒素常常要延迟1~3刻钟后才开始;
(2)激素去除后,效应很快消失,而霍乱毒素在去除后生物效应还可持续几小时到几天;
(3)一种激素的受体只在少数几种细胞中存在,而霍乱毒素的受体在很多细胞膜上都有分布。
凝集素受体凝集素是从各种植物或低等动物组织中分离而得到的一类特殊蛋白质,能与动植物细胞表面的受体发生特异性结合产生一系列的生理效应。它们通过与细胞表面寡糖结构决定簇的交互作用导致细胞发生凝集,因而又称凝集素。最早发现它们能诱导红细胞发生凝集而用于临床血型分类,故又有植物血凝素之称。目前已报道的凝集素多达500种。
与凝集素特异结合的受体位于细胞表面,具有复杂的寡糖链结构,它们拥有特殊的联结、侧链及多肽附着物。在这些寡糖链间还存在非共价键(如氢键)的相互作用。凝集素与受体寡糖链的结合部位以往都认为是末端的糖残基,现在证明还能与多糖核心部位相结合。
不同的凝集素可以各有专一的受体,但也可以具有共同的受体。总的来讲,凝集素受体的专一性比抗原-抗体的专一性要差。
凝集素使细胞凝集是由于一个凝集分子能与细胞表面两个或两个以上的结合点结合后所引起的。
当凝集素与其专一受体结合后可引发细胞多种生理效应,如细胞凝集,特别是肿瘤或转化细胞可发生高效价的凝集作用;诱发淋巴细胞发生丝裂现象以及对膜透性、受精过程、吞噬及细胞增殖等都可产生一定的影响。
- 参考书目
- F.B.Finean,R.Coleman,R.H.Michell,Membranesand Their Cellular Functions,2nd ed., Blackwell Scienlific Pub.,Oxford,1978.