[拼音]:tiwen tiaojie
[外文]:thermoregulation
动物保持体温相对恒定的能力,是动物在长期进化过程中获得的较高级的调节功能。体温即机体的温度,通常指身体深部的温度。
变温动物和恒温动物
较低等的脊椎动物如爬行类、两栖类和鱼类,以及无脊椎动物其体温随环境温度而改变,不能保持相对恒定,这些动物叫做变温动物或冷血动物。变温动物对环境温度变化的适应能力较差,到了寒季,其体温降低,各种生理活动也都降至极低的水平。进化至较高等的脊椎动物如鸟纲和哺乳纲动物,逐渐发展了体温调节功能,能够在不同温度的环境中保持体温的相对恒定,这些动物叫做恒温动物或温血动物。还有一些哺乳动物如刺猬等,则介乎上两类动物之间。在暖季,体温能保持相对恒定;到了寒季则体温降低,蛰伏而冬眠。
体温的行为调节和自主调节
体温调节的方式有两类:
(1)行为性体温调节,动物通过其行为使体温不致过高或过低的调节过程。如低等动物蜥蜴从阴凉处至阳光下来回爬动以尽量减小体温变动的幅度。人在严寒中原地踏步、跑动以取暖,均属此种调节。人类能根据环境温度不同而增减衣著,创设人工气候环境以祛暑御寒,则可视为更复杂的行为调节。
(2)自主性体温调节,动物通过调节其产热和散热的生理活动,如寒颤、发汗、血管舒缩等,以保持体温相对恒定的调节过程。
产热、散热过程及其动态平衡
产热过程机体代谢过程中释放的能量,只有20~25%用于作功,其余都以热能形式发散体外。产热最多的器官是内脏(尤其是肝脏)和骨骼肌。静息时肝、肾、胃肠等内脏器官的产热量约占机体总产热量的52%;骨骼肌产热量约占25%。运动时,肌肉产热量剧增,可达总热量的90%以上。冷环境刺激可引起骨骼肌的寒颤反应, 使产热量增加4~5倍。产热过程主要受交感-肾上腺系统及甲状腺激素等因子的控制。因热能来自物质代谢的化学反应,所以产热过程又叫化学性体温调节。
散热过程体表皮肤可通过辐射、传导和对流以及蒸发等物理方式散热,所以散热过程又叫物理性体温调节。辐射是将热能以热射线(红外线)的形式传递给外界较冷的物体;传导是将热能直接传递给与身体接触的较冷物体;对流是将热能传递给同体表接触的较冷空气层使其受热膨胀而上升,与周围的较冷空气相对流动而散热。空气流速越快则散热越多。这 3种形式发散的热量约占总散热量的75%,其中以辐射散热最多,占总散热量的60%。散热的速度主要取决于皮肤与环境之间的温度差。皮肤温度越高或环境温度越低,则散热越快。当环境温度与皮肤温度接近或相等时,上述3种散热方式便无效。如环境温度高于皮肤温度,则机体反而要从环境中吸热。变温动物即常从环境中获得热能。
皮肤温度决定于皮肤的血流量和血液温度。皮肤血流量主要受交感-肾上腺系统的调节。交感神经兴奋使皮肤血管收缩、血流量减少,皮肤温度因而降低。反之,则皮肤血管舒张,皮肤温度即行升高。所以说皮肤血管的舒张、收缩是重要的体温调节形式。
蒸发是很有效的散热方式。每克水蒸发时可以吸收0.58千卡的汽化热。常温下体内水分经机体表层透出而蒸发掉的水分叫做无感蒸发。其量每天约为1000毫升。其中通过皮肤的约600~800毫升;通过肺和呼吸道的约200~400毫升。一般在环境气温升到25~30℃时,汗腺即开始分泌汗液,叫做出汗或显汗──可感蒸发。环境气温等于或高于体温时,汗和水分的蒸发即成为唯一的散热方式。出汗是人类和有汗腺动物在热环境中主要的散热反应。无汗腺的动物如狗等,主要以热喘及流涎等方式来增加蒸发散热。汗腺分小汗腺和大汗腺两种:小汗腺分布于人体全身皮肤,以手掌、足跖和前额最密。猴、猫、鼠等的汗腺主要分布于足跖部,它受交感神经的胆碱能纤维支配。大汗腺开口于毛囊的根部,分布于动物全身皮肤,而人类则较不发达,局限于腋窝、外阴部等处,它受肾上腺素能纤维支配。出汗反射也分两类:由温热刺激引起的为温热性发汗。此种发汗见于全身,而以躯干部最多,额面部次之。其主要中枢在下丘脑前部。由精神紧张或疼痛引起的为精神性发汗,主要见于手掌、足跖等处,不属于散热效应。一般认为其中枢在大脑皮层的运动前区。
产热和散热的动态平衡体温的稳定决定于产热过程和散热过程的平衡。如产热量大于散热量时,体温将升高;反之,则降低。由于机体的活动和环境温度的经常变动,产热过程和散热过程间的平衡也就不断地被打破,经过自主性的反馈调节又可达到新的平衡。这种动态的平衡使体温波动于狭小的正常范围内,保持着相对的稳定。
人体正常体温及其生理变动
临床上一般采取从腋窝(人工体腔)、口腔和直肠内测量体温的方法。正常人体的直肠温度平均约为37.3℃,接近于深部的血液温度;口腔温比直肠温低0.2~0.3℃,平均约为37.0℃;腋窝温比口腔温又低0.3~0.5℃,平均约为36.7℃。正常生理情况下,体温可随昼夜、年龄、性别、活动情况不同而有一定的波动。一昼夜中,清晨2~4时体温最低,午后4~6时最高,变动幅度不超过1℃。这种近日节律并不因生活习惯的变动而改变,它很可能与地球的自转周期有关。新生儿的体温略高于成年人,老年人则稍低于成年人。婴儿的体温调节机能尚未完善,可受环境温度、活动情况或疾病的影响而有较大的波动。女人在月经来潮时体温可上升约0.2℃,至排卵日(经后第14天)又再上升0.2℃左右。 这可能是雌激素的作用。排卵后体温逐渐下降至经前水平,这是孕激素的影响,临床上常据此来了解妇女是否排卵。剧烈的肌肉运动、精神紧张或情绪刺激也可使体温升高1~2℃。在酷热或严寒环境中暴露数小时,体温可上升或下降1~2℃。
机体对温度变化的感受
周围环境的温度变化,可改变体表温度而刺激皮肤的冷、热感受器,引起传入冲动的发放。皮肤温度感受器呈点状分布,其中冷点较多,约为热点的4~10倍。 冷感受器的放电频率远远高于热感受器。通常认为皮肤对寒冷刺激比较敏感。腹腔内也有热感受器,其传入纤维在内脏大神经中。
用变温管来加温或冷却动物脑和脊髓的局部组织,证明脑和脊髓都有温度感受神经元存在,而以下丘脑的最为重要。提升狗的视前区-下丘脑前部的温度,可引起代谢率下降、皮肤血管舒张、皮肤温度升高和热喘等效应。降低该部位的温度,则出现相反的效应。用微电极记录神经元单位放电,结果看到在狗的视前区-下丘脑前部神经元中,30%为对脑温升高发生放电效应的热敏神经元,其Q10较高;10%为对脑温降低发生反应的冷敏神经元,其Q10较低。其余60%对脑温改变不敏感。但有资料表明,来自皮肤温度感受器的传入信息可以引起其中一些神经元的放电。这说明视前区-下丘脑前部能接受和整合来自中枢和外周的温度觉信息。
体温调节中枢
体温调节的基本中枢在下丘脑。切除下丘脑以上的前脑的动物即“下丘脑动物”,仍能保持接近正常的体温调节功能。而切除中脑以上的全部前脑(包括下丘脑)的动物则不能保持体温的相对稳定。用局部加热或电刺激猫的下丘脑的前部,可引起热喘、血管舒张和足跖发汗等散热效应。破坏该区后,猫在热环境中的散热反应能力丧失,但对冷环境的反应(寒颤、竖毛、血管收缩、 代谢率升高等)仍存在。破坏下丘脑后部内侧区的效果,则正相反,对冷环境的反应丧失。传统生理学据此认为,在下丘脑前部存在着散热中枢,而下丘脑后部则存在着产热中枢。两个中枢之间有着交互抑制的关系,从而保持了体温的相对稳定。
在下丘脑前部还存在着发汗中枢。下丘脑后部内侧区存在着寒颤中枢,它对血液温度变化并不敏感,但对来自皮肤冷觉感受器的传入信息比较敏感。电刺激下丘脑前部(散热中枢)可以抑制寒颤;冷却视前区-下丘脑前部则可以引起寒颤。这表明下丘脑前部有冲动输入至下丘脑后部。
下丘脑与体温的行为调节亦有关。对鼠猴进行训练,使它每次从冷室返回时能自行拧开热气开关取暖。如此时突然将其视前区-下丘脑前部的温度由36℃提高到42℃,它就立即关闭热气而打开冷气。这表明体温的行为调节受下丘脑的控制,而体温调节中枢对体内外温度变化的反应,则取决于大脑对来自外周和中枢的多种温度觉信息整合的结果。
体温调节反射与控制论
体温的自主性调节主要通过反射来实现。环境温度或机体活动的改变将引起体表温度或深部血温的变动,从而刺激了外周或中枢的温度感受器。温度感受器的传入冲动经下丘脑整合后,中枢便发出冲动(或引起垂体释放激素),使内分泌腺、内脏、骨骼肌、皮肤血管和汗腺等效应器的活动发生改变,结果调整了机体的产热过程和散热过程,从而可以保持体温的相对稳定(见温度觉)。
这一调节机制,与控制论的原理颇为符合(见图)。
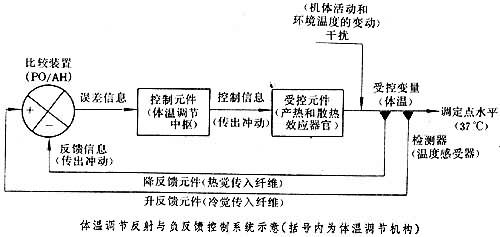
体温调节机制相当于负反馈控制系统。当体温高于或低于37℃(偏离了“调定点”水平)时,温度感受器的传入冲动(反馈信息)经视前区-下丘脑前部(比较装置)整合后,中枢传出冲动便调整了产热和散热器官的活动,使体温复归原初的水平。在自动控制系统中,输入比较装置以决定“调定点”水平的参考信号是由操纵者事先给定的。但是在体温调节机制中,目前还没有证据足以表明中枢神经系统的哪一部分会发出这样的参考信号。较为普遍的看法认为:由机体各部传入的无数的温度觉信号,在下丘脑进行比较和整合后,决定体温的调定点水平(见生物控制论)。
体温调节的调定点假说
认为在下丘脑的前部存在着高Q10的热敏神经元和低Q10的冷敏神经元。这两种温度感受神经元的活动共同作用的结果,决定了体温调节的调定点。体温偏离这一调定点水平时,可通过反馈系统的调节,使体温回到调定点水平。调定点是可以变动的。发热就可能是由于细菌内毒素等致热原使视前区-下丘脑前部的热敏神经元阈值升高、调定点水平上移所致。致热原的作用可能是通过前列腺素 E这一中间环节,而阿斯匹林能够抑制前列腺素的合成,因而起退热的作用。体温的一些节律性变化,许多学者亦用调定点的节律性变动来解释。
有些学者认为视前区-下丘脑前部温度感受神经元的活动决定了调定点,而下丘脑后部则为传出神经元发出的部位。另一些学者认为视前区-下丘脑前部感受脑温的变化,而下丘脑后部则对来自皮肤的大量温度觉(主要是冷觉)信息和从视前区-下丘脑前部传来的温度觉(主要是热觉)信息,进行整合并决定调定点水平。调定点假说的根据尚不够充分,争论也较多,尚待探索。
中枢递质与体温调节
下丘脑体温调节中枢含有丰富的单胺能神经元,释放去甲肾上腺素(NE)、5-羟色胺(5-HT)和多巴胺(DA)等神经递质。灌流动物侧脑室或下丘脑的实验证明:5-HT可引起猫、狗、猴的体温升高,并伴有寒颤和外周血管收缩反应,但兔和大鼠对5-HT的效应则与此相反。去甲肾上腺素可引起猫、狗、牡牛的体温降低,并伴有外周血管舒张等效应,但兔、羊、大鼠的效应相反。多巴胺的作用与去甲肾上腺素大致相似。将动物置于冷或热环境中,也能相应地引起脑内释放这类递质。上述研究表明,中枢递质可能在体温调节中起重要作用。
体温调节对环境的适应
人体进入高温或低温环境时,各项生理活动均有显著的变化。这些变化实质上反映了机体在调整它的各种机能以适应环境温度的剧烈变动。在低温或高温地区居住较久的人,可对环境温度产生顺应。此时对环境温度的耐受性增强,功能调节的效率提高。例如对热顺应的人在温热环境中蒸发散热的效率较高,而且血浆量增多,随尿和汗液排泄掉的盐减少。对冷顺应的人,开始寒颤的温度(阈值)较低,基础代谢率增高。
人类对环境温度改变所产生的自主性体温调节是很有限的,更多的是行为性调节。人类能够改造环境,使其更适于人类的生存。这是主动性适应的能力。