[拼音]:chaoxunhuan lilun
[外文]:hypercycle theory
研究分子自组织的一种理论。大分子集团借助于超循环的组织形成稳定的结构,并能进化变异。这种组织也是耗散结构的一种形式。超循环是较高等级的循环,指的是由循环组成的循环。在大分子中具体指催化功能的超循环,即经过循环联系把自催化或自复制单元等循环连接起来的系统。从动力学性质看,催化功能的超循环是二次或更高次的超循环。超循环理论可用以研究生物分子信息的起源和进化,并可用唯象的数学模型来描述。超循环理论是联邦德国生物物理学家M.艾根在1971年提出的。曾有不少学者提出各种理论来研究生物信息的起源和进化。艾根总结了大量的生物学实验事实,提出了超循环理论。
原理生命的发展过程分为化学进化和生物学进化两个阶段。在化学进化阶段中,无机分子逐渐形成简单的有机分子。在生物学进化阶段中,原核生物逐渐发展为真核生物,单细胞生物逐渐发展为多细胞生物,简单低级的生物逐渐发展为高级复杂的生物。生物的进化依赖遗传和变异,遗传和变异过程中最重要的两类生物大分子是核酸和蛋白质。各种生物的核酸和蛋白质的代谢有许多共同点,所有生物都使用统一的遗传密码和基本上一致的译码方法,而译码过程的实现又需要几百种分子的配合。在生命起源过程中,这几百种分子不可能一起形成并严密地组织起来。因此,在化学进化阶段和生物学进化阶段之间有一个生物大分子的自组织阶段,这种分子自组织的形式是超循环。如核酸是自复制的模板,但核酸序列的自复制过程往往不是直接进行的。核酸通过它所编码的蛋白质去影响另一段核酸的自复制。这种结构便是一种超循环结构。这种大分子结构是相对稳定的,能够积累、保持和处理遗传信息。另一方面,这种结构在处理遗传信息时又会有微小的变异,这又成为生物分子发展进化的机制。为了根据生物大分子自组织的基本要求建立生物进化变异模型,艾根提出一组唯象的数学方程,并得到一些具有启发意义的结果。选择的对象不是单一的分子种,而是拟种,即以一定的概率分布组织起来的一些关系密切的分子种的组合。信息选择的积累以自复制子单元最大信息容量为上限,超过这个限制就不能保证拟种的内部稳定性。可以认为,拟种的内部稳定性是进化行为更本质的属性。考虑到生物体内进行着许多必不可少的生化反应,需要许多不同的蛋白质和核酸参加,它们总的信息量远大于已知的最精确复制机制所允许的最大信息容量。这一实验事实表明,只有经过超循环形式的联系才能把自复制和选择上稳定的单元结合为较高的组织形式,以便下一步再产生选择上稳定的行为。超循环有如下一些重要性质:
(1)超循环使借助于循环联系起来的所有种稳定共存,允许它们相干地增长,并与不属于此循环的复制单元竞争。
(2)超循环可以放大或缩小,只要这种改变具有选择的优势。
(3)超循环一旦出现便可稳定地保持下去。总之,生物大分子的形成和进化的逐步发展过程需要超循环的组织形式。它既是稳定的又允许变异,因而导致普适密码的建立,并在密码的基础上构成细胞。
具有超循环结构的生物大分子的进化可用一组微分方程来描述。设有n个物种,其状态变量记作xi(i=1,2,...,n),它们形成自复制的催化超循环结构需要依靠外界的能量和物质流来维持,在不断复制过程中会产生误差,这样就导致优势物种的变异。描述这种进化过程的方程组是
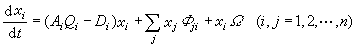
式中Ai是复制率,Qi是模板的品质因子,Di是分解率,Фji是j物种到i物种的误合成系数,Ω是环境影响因子,可以是非线性的。因此在这一方程组中包含了物种变异的各种因素。对这个方程进行分析、数值计算或定性讨论,就可以得出有关物种进化的各种趋势。例如选择不同的误合成系数Фji可以得到不同的结果。当误合成系数Фji很小时,在初始时刻某一物种占优势的状态便逐渐变异到终了时刻另一物种占优势的状态。如果误合成系数相当大,最终便形成多种物种并存的状态。这一方程组及其所得结果,使人们有可能用数学工具讨论进化过程。
理论意义超循环理论对于生物大分子的形成和进化提供了一种模型。对于具有大量信息并能遗传复制和变异进化的生物分子,其结构必然是十分复杂的。超循环结构便是携带信息并进行处理的一种基本形式。这种从生物分子中概括出来的超循环模型对于一般复杂系统的分析具有重要的启示。如在复杂系统中信息量的积累和提取不可能在一个单一的不可逆过程中完成,多个不可逆过程或循环过程将是高度自组织系统的结构方式之一。超循环理论已成为系统学的一个组成部分,对研究系统演化规律、系统自组织方式以及对复杂系统的处理都有深刻的影响。
- 参考书目
- M.Eigen,P.Schuster, The Hypercycle,Springer-Verlag, Berlin,1979.