[拼音]:danshui shengtai
[外文]:freshwater ecology
研究内陆水体中生物与生物之间以及生物与环境之间的相互关系。包括生物个体、种群、生物群落和生态系统等层次,而生态系统已成为目前的研究重心。内陆水体包括江河、湖泊、水库、池塘等,具有广泛的地理分布和悬殊的大小规模,其总面积约占地球面积的0.5%。这些水体不仅是人类生活与工农业生产用水的主要来源,而且在航运、发电、渔业等方面给人类带来了许多利益。同时,内陆水体作为生物圈的组成部分,对调节全球气候、维持自然环境的稳定性起着重要的作用。淡水生态系统的研究,有助于阐明系统中的物质循环、能流以及演替和平衡的规律,为加强内陆水体的水质管理、防治水体污染或富营养化、合理开发和利用水体生物生产力提供科学依据。
淡水生态系统
指一定水域内所有生物(即生物群落)与它们的理化环境相互作用,通过物质循环和能流而构成的具有一定结构和功能的统一体 (图1)。
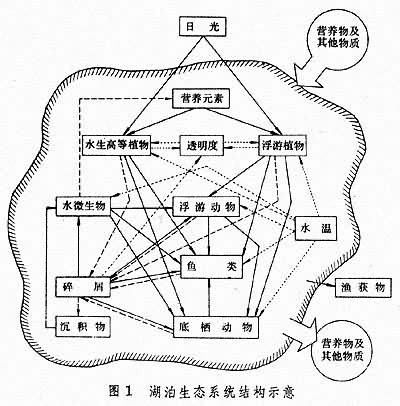
生态系统的非生物组分包括:气候条件(温度、光照及其他物理因素),参加物质循环的无机物质(碳、氮、磷、二氧化碳、水等),以及联系生物和非生物的有机化合物(蛋白质、碳水化合物、脂类、腐殖质等)。生态系统的生物组分依其生态功能可分为:生产者,主要指浮游植物和水生高等植物;大型消费者,包括浮游动物、底栖动物、游泳动物等;以及微型消费者,主要是细菌和真菌。淡水生态系统的结构,不仅视静水与流水两大类型而异,而且同一类型的各种栖息地之间往往也有明显的差别。
生态系统的生物组分通常分布在空间的不同层次,它们之间主要以食物链(网)的形式相互联系。在生态系统的代谢过程中,绿色植物以日光为能源将无机物质合成可供消费者利用的有机物质,其食物潜能沿着不同的食物链逐级传递;死亡有机物质则被微型消费者分解,结果释放出可被绿色植物重新利用的无机营养物。于是,生态系统中不断地进行着营养物质在环境与生物之间的往复循环,以及能量从日光到生物的单向流动。物质循环和能流是生态系统的最重要的功能,而生物生产力是生态系统的这两大功能的综合表征。
任何生态系统都是由简单到复杂,从低级到高级不断发展的。淡水水体发育和演替的基本模式,是从贫营养到富营养和由水体到陆地。生态系统的发育,一方面起因于系统内生物组分的活动,另一方面受外界因素的影响。在经过一定发育阶段的生态系统中,各生物组分之间、群落与环境之间、以及结构与功能之间的相互关系是相对稳定和协调的,可通过信息反馈来维持自身的动态平衡。但生态系统的自我调节能力有限,其平衡状态往往因强度过大的外来干扰而遭到破坏。20世纪60年代以来,淡水水体的严重污染或富营养化,都是人为因素所造成的自然生态平衡失调的实例。因此,加强淡水生态系统的管理,使之朝着有利于人类的方向发展,已成为关系到国计民生的一个重大问题。
淡水环境
地球上的水量共约137×108立方公里,其中96.5%的水积蓄在海洋,余下的分布在两级冰冠、冰川、地下、内陆水体、土壤和大气中。内陆水体的水量仅占地球总水量的0.016%,其中淡水水体的水量占55%,盐湖和内陆海的水量占45%。地球各部分的水量分布,是通过降水、径流和蒸发所构成的水循环而维持相对稳定的,内陆水体在这一循环过程中起重要作用。
内陆水体大体上可分为静水水体(如湖泊、水库、池塘)和流水水体 (包括泉、溪流及江河)两大类。这些水体不仅地理位置和气候条件不同,而且具有多种多样的形态、结构和化学组成,因此生物的环境条件极为复杂。在淡水生态学研究中,水环境的温度、光照(决定于透明度)、水流(尤其在河流中)、溶解气体(氧和二氧化碳)、营养盐类(主要是磷酸盐和硝酸盐)等,通常被认为是最重要的限制因素。
湖泊
静水水体的主要类型。地球上可利用的淡水大部分贮存在湖泊中。从地质年代来看,湖泊的存在是短暂的,现有湖泊大都形成于较晚的历史时期。如北欧、加拿大和美国北部地区的很多湖泊,大约在1~1.2万年前最后的冰川退却时形成。按照湖盆的成因,湖泊可分为构造湖、冰川湖、火山湖、山崩湖、溶解湖、河成湖、风成湖、海湾湖等类型。不同类型的湖泊具有不同的底质和形态特征,而这些特征直接关系到湖泊的理化环境。
湖泊作为生物的栖息地,通常可按光照条件分为沿岸带、敞水带和深水带 3个不同的生境。沿岸带指靠近湖岸的浅水区,日光可以直射到底,一般被水底长出的水生高等植物所占据。沿岸带以外从水面直到光的有效透射深度(即补偿深度)为止的水层称敞水带。在所谓补偿深度处光合作用放出的氧气正好满足呼吸作用的消耗,其光照强度通常为饱和光强度的1%。上述两个区域都属于光亮带。敞水带以下,即由补偿深度直至水底的区域是深水带,这里光照微弱或无光线,不见大型植物分布,但底部可有大量微生物滋生。浅水湖泊不存在深水带,其他湖泊的敞水带和深水带都大于沿岸带。一般说,湖泊的沿岸带光照、氧气和营养条件最佳,生物的种类和数量都很丰富。因此在水体生产力研究中常用湖岸线发育指数(DL)作为湖泊生产性能的一个指标。
湖泊中温度(热)、溶解气体和营养盐类的空间分布主要取决于湖水的运动,由于引起湖水运动的主要因素(风和水的密度差)直接受气候影响,因此湖中温度、光照、养分等要素空间分布的季节变化随着湖泊的地理位置的不同而有很大差别。在温带地区,夏季的风力只能引起湖泊上层的水循环(湍流混合),致使湖中(除浅水湖泊外)出现温度随深度下降的正向分层现象。按照湖中的温度梯度可将湖水分为湖上层、温跃层和湖下层,其中温跃层的温度随深度急剧下降(温度通常每米可下降1℃)。冬季湖面结冰,冰层阻止湖中热量的进一步散失,结果出现湖下层温度高于湖上层的反向分层现象。春季和秋季分别是湖泊吸热和放热的时期,在强大的湍流混合和水密度差所引起的对流混合作用下,湖泊的上下水层出现循环(翻转)而处于同温状态。热带湖泊温度的季节变化很小,仅在某些年份于较冷的季节出现水循环,一般情况下保持着温度梯度很小的正分层。根据湖泊的热学特征,可将世界上的大多数湖泊分为6个类型:二次混合湖(温带,水温4℃左右);冷一次混合湖(两极,水温不超过4℃, 夏季循环);热一次混合湖(温带或亚热带,水温不低于4℃,冬季循环);寡混合湖(热带、很少混合);多次混合湖(赤道、高山地区,连续混合);和部分混合湖(由盐类引起的永久性化学分层,水循环仅限于湖上层)。伴随湖泊的热分层,也出现氧分层现象。在温带湖泊夏季热分层期间,湖中的温跃层往往位于补偿深度之下,结果湖下层因氧供应完全断绝而陷于缺氧状态,这种情况在富营养湖泊中更为严重。
湖泊的化学组成和营养状况,因湖盆及其流域的地理和地貌特征以及湖泊的发育过程而异。目前广泛采用的世界湖泊分类方法有两种:一是以生产力为依据的正常湖泊的贫营养-富营养系列;二是具有异常化学性质的特殊湖泊类型。湖泊的生产力或“肥力”取决于外源营养物、地质年龄和深度,大多数湖泊都可按初级生产力分类。贫营养湖泊一般较深,养分含量和初级生产力均低,处于地质学上的年轻阶段。相反,富营养湖泊较浅,营养物含量和初级生产力均高,并且经历了较长的地质年代。因此,在比较水色、透明度、底质、溶解氧、pH、无机和有机物含量,以及生物的组成和多度时,典型的贫营养湖泊与富营养湖泊往往是完全不同的。在贫营养-富营养系列中,按照给定指标的梯度,可以划分出许多中间型湖泊,如贫-中营养湖、中营养湖、中-富营养湖等。特殊类型的湖泊主要包括:腐殖营养湖 (含高浓度腐殖酸),盐湖(含盐量高,分布于干旱气候地区)和酸性或碱性湖(分布于活动的火山地区,受岩浆的影响大)。在这些湖泊中,化学条件严酷,通常仅有很少的具有特殊适应能力的生物。
水库
近200年来,世界各地所建水库的贮水量在内陆水体中占有相当的比重。水库的环境条件与天然湖泊有许多相似之处,上述有关天然湖泊的生境划分和营养分类基本上适用于水库。水库的特点是水位不稳定和混浊度大,所以生物生产力往往低于天然湖泊。水库的热量、氧气和营养物收支依其排水方式而异。如果水闸设在水坝底部,那么排出的就是温度低、营养物丰富和氧气贫乏的水,而温度高、营养物缺乏和氧气充足的水留在水库中。这不仅会使水库失去必需的营养物,从而导致生产力降低,而且将对下游地区产生多方面的影响。若从水库表面排水,其结果则与天然湖泊相同。
池塘
小型静水水体,沿岸带较大,而敞水带和深水带都较小或不存在, 故水体中的理化条件较为一致。一部分池塘是自然形成的(天然池塘,如河流改道后留下的部分旧河床、石灰岩底质被溶解所形成的洼地),或产生于某些动物(如河狸)的活动,但数量最多的是为了工农业用水而修筑的人工池塘。池塘的水量变动较大,甚至有些池塘在一年中的部分时间内是干涸的,这类池塘常称临时池塘。多数池塘都很肥沃,尤其是作为渔业水体的池塘,往往经人工施肥,因此常有较高的生产力。
河流
流水水体的主要类型,其贮水量约占内陆水体总水量的 0.5%, 亦是地球上水循环的一个重要环节。与静水水体不同,河流的首要特征是具有连续的水流,这在很大程度上决定着河流中许多理化成分和小型生物的分布。河水的流速决定于河床的坡降、深度、宽度和粗糙度,并且在很大程度上受排水量的影响。同一河流的不同部分和不同时期的流速常有很大的变化;在特定条件下,河流的某些区域可能处于静水状态。因此,河流中通常存在急流带和滞水带两个不同的生境:前者指浅水区,流速大,底质坚硬;后者指深水区,水流缓慢,底质常松软。某些河流的下游水流相当平缓,急流带与滞水带的差别基本消失,这部分河段常被称为河道带。河流的另一个特征是深度和横断面显著小于湖泊和水库,具有较大的水-陆连接面,成为更开放的生态系统。实际上,大多数河流中有机物质主要来自陆地和邻近的静水水体,尤其是处于森林地带和肥沃平原的河流接受外源物质更多。由于河流的深度小,与空气的接触面大,河水又不停地运动和混合,河流中的溶解氧通常较充足,并且几乎没有热分层或化学分层现象。河流排水量的季节变动大,不同河段的流量差异显著,外源物质的输入具不确定性,所以河流的化学组成往往随时间和距离而有很大的变化。同时,河流往往被看作有效的废物处理系统,因而容易遭受有毒物质污染。
泉
地球上数量很少的一类小型流水水体,具有相对稳定的温度、流速和化学组成。常见的几种较为重要的泉是:温泉(分布于火山地区,通常含盐量高)、硬水泉(分布于石灰岩地区,具有所在地的平均温度)和软水泉(水从页岩、沙岩和石英岩渗出)。泉流的理化性质固然在泉水流向下游的过程中发生变化,但是任一给定点的理化性质总是比较稳定的,因此提供了一个天然的生态梯度。鉴于某些大泉流中群落结构和代谢的稳定性,许多生态学家都把泉看作一个重要的研究领域。
淡水生物群落
生活在淡水栖息地的各类生物,通常分布在不同的空间层次和占据着各自的生态位,彼此之间存在着复杂的相互关系,这些生物种群的集合体被称为生物群落。在生态学研究中,除了按水体或生境来研究整个生物群落外,还可以着眼于一些规模较小的群落单元,如浮游生物群落,底栖生物群落、周丛生物群落等。
淡水生物的生态类别
淡水生物依其在群落中所处的营养级可以分为:生产者(自养生物),主要包括浮游植物、水生高等植物和着生藻类,其次是一些光合细菌和化能自养细菌;大型消费者(牧食性和捕食性异养生物),指吃其他生物或有机碎屑的动物,主要有浮游动物、底栖动物和各种鱼类;微型消费者(腐食性异养生物或分解者),主要指细菌和真菌。淡水生物群落的每一个营养级上分布着多种生物,它们在群落代谢中所起的作用很不相同,因此常有优势种类与次要种类的区别。淡水生物依其生活方式或生活习性,可分为浮游生物、游泳动物、底栖生物、周丛生物和漂浮生物5个生态类群。
静水生物群落
按照静水栖息地的3个生境类型,静水生物群落可分为沿岸带群落、敞水带群落和深水带群落,这些群落的相对重要性主要决定于水体中3个生境的相对大小。一般说,沿岸带是池塘的主要生产区,而湖泊和水库的主要生产区则是敞水带。
沿岸带群落沿岸带的生产者主要包括底生植物和各种藻类,其次是漂浮植物。底生植物的分布通常有明显的水平成带现象,即从岸边到深处随着水深的变化而出现3个不同的植物带:
(1)挺水植物带,植物的茎叶高出水面,主要包括芦苇、水烛、阔叶菰、莲等;
(2)浮叶植物带,植物的叶浮在水面,主要有菱、荇菜、睡莲等;
(3)沉水植物带,植物体完全沉没在水下,主要种类为眼子菜科植物,其次有苦草、黑藻、等。藻类主要包括硅藻、绿藻和蓝藻。除了一部分丝状藻类和着生藻类依附于底生植物外,大部分藻类都为浮游性。底生植物的分布主要决定于水层的透明度,其生长情况常与浮游藻类的生长呈负相关。在富营养化水体中,由于蓝藻水华严重阻碍了光的透射,底生植物往往濒于灭绝。漂浮植物主要有浮萍、满江红、水浮莲等,在水面平静的地方(如池塘和湖湾)常有大量分布。
沿岸带的消费者生物极为丰富,属于5个生态类群的各级消费者都有存在。某些动物(尤其是周丛生物)的水平成带分布常与底生植物的分布相平行,但有很多种类几乎分布在整个沿岸带,并且动物的垂直成带现象比水平成带更为显著。周丛生物主要有螺类、某些昆虫幼虫、原生动物、水螅、轮虫、各种蠕虫、苔藓虫等。底栖动物中以多种昆虫幼虫、环节动物(寡毛类)和软体动物(螺、蚌)占优势。浮游动物主要有大型的枝角类、桡足类的某些种类、介形类和轮虫。很多甲壳动物都有笨重的外壳,常采用爬行方式运动,这是与敞水带的种类明显不同的特征。漂浮动物通常仅有几种水面昆虫,如鼓甲、黾蝽等。游泳动物主要有多种昆虫(幼虫和成虫)、两栖类(蛙)、爬行类(龟、蛇)和各种鱼类。其中鱼类来往于沿岸带与敞水带之间,常在沿岸带觅食和繁殖。
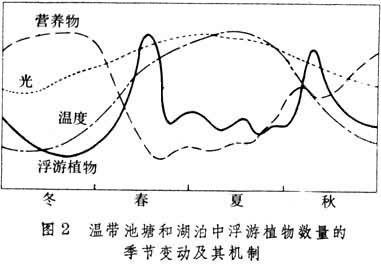
敞水带群落生产者包括浮游植物和一些浮游自养菌。浮游植物主要有甲藻、硅藻、绿藻和蓝藻,个体小,但生产力相当高。在北温带湖泊中,随着温度、光照和营养物再生的季节变化,浮游植物于春秋两季各出现一次高峰(水华),夏冬两季则处于较低水平(图2)。春季水华主要由硅藻所引起,而秋季水华通常归因于蓝藻。
消费者主要包括浮游动物和各种鱼类。浮游动物主要为桡足类、枝角类和轮虫,它们与沿岸带的那些种类有明显区别。很多浮游动物属滤食动物,但也有不少捕食性种类。浮游动物的种类组成和数量分布随浮游植物而变动,往往在浮游植物高峰之后出现浮游动物高峰。敞水带的鱼类通常与沿岸带的相同,但大型水体中可能有某些鱼类局限于敞水带。这些鱼类分布于不同水层,一般都有很大的活动范围。在中国人工经营的水体中,以浮游生物为食的中上层鱼类(鲢和鳙)已成为优势种群。
深水带群落深水带不存在生产者,消费者种类也不多,其食物供应依赖于沿岸带和敞水带。群落的主要成员是细菌和真菌,它们所提供的再生营养物常通过水流和游泳动物带到其他区域。大型消费者主要有摇蚊幼虫、颤蚓、球蚬和幽蚊幼虫。前3种属于底栖动物,最后1种是部分浮游动物,这些动物都有适应缺氧环境的能力。
流水生物群落
流水生物群落可按河流所在地的3类生境分为急流带群落、滞水带群落及河道带群落。河流上下游的环境条件差别显著,河流生物的分布有明显的纵向成带现象,这点与静水生物的水平成带分布不同。
急流带群落由典型的河流生物组成,主要是一些着生藻类和各种昆虫幼虫。生活在这里的动物都具有特化的形态结构,明显地适用于流水环境。平常见到的浮游生物来自邻近的静水水体。
滞水带群落生产者有丝状藻类和一些沉水植物。消费者主要包括一些穴居或底埋动物和多种鱼类,后者为了觅食亦常在急流带出现。在某些流速很低的河流中还生长着浮游生物。
河道带群落其结构类似于静水生物群落。除河流生物外还可见到很多静水生物。但由于河床底质的不均匀性,底栖动物通常以成团的形式分布。
养分循环和能流
养分循环
淡水生态系统是生物圈的一个组成部分,一方面与大气保持着连续的气体交换,另一方面通过径流与陆地、农田和邻近的其他水体存在着广泛的联系。因此,淡水生态系统中的养分处于经常的变动状态,其输入与输出关系依水体的类型、形态结构和地理分布而异。大多数静水水体排出的是养分贫乏的表面水,所以养分的输出明显地小于输入,水体有向富营养化发展的趋势。
虽然水体可由多种途径接受一定的外源养分,但是经常的和大量的养分供应主要依赖于系统内部的循环。这一循环的基本过程是:绿色植物以日光为能源,由无机养分合成有机物质,供消费者利用,同时通过生物的呼吸和死亡有机物质的分解,养分重新释放出来。
淡水生态系统中有机物质的分解,是生物的和非生物的因素共同作用的结果,但是一般起主要作用的是以这些有机物质为食物来源的腐食者生物。水体中的有机物质,一部分是系统内的动植物尸体和排泄物,另一部分是来自其他系统(外源)的有机物质,包括人类排放的污水和施入的有机肥料。在有机物质分解的过程中,各种异养微生物通常是同时或交替起作用的,任何一个分解者种群都不能单独实现把有机物质完全分解的任务。现知,一些小型的腐食性或碎屑食性动物,如原生动物、线虫、介形类等,在有机物质分解过程中所起的作用很大。在某些生态系统中,小型动物对有机物质的分解甚至比细菌或真菌更为重要。
微生物的分解过程只是营养物再生的途径之一,而且在某些情况下还处于从属地位。例如,在浮游生物占优势而以牧食食物链为主要能流途径的生态系统中,直接从浮游动物的排泄物和分泌物中返回的养分,常比经微生物分解所得到的量高得多。研究表明,浮游动物的排泄物中含有许多可溶性的无机和有机化合物,这些养分能直接被浮游植物所利用。细菌在分解植物组织时会将所获得的磷酸盐和硝酸盐保留在体内,只有当它们被食碎屑的动物消费后营养物才能释放出来。
在淡水生态系统中,养分的循环过程不是均衡的,而是存在着养分的吸收与释放的周期变化。经32磷示踪试验发现,湖泊中大部分的磷或存在于生物体内,或被结合在沉积物中,而水中的溶解磷至多只有10%。因此,沉积物中养分的释放率,在很大的程度上反映了养分循环的速度。由于不同水体的形态结构和理化状况不同,沉积物中养分的释放率往往差别很大。一般情况下,小型浅水湖泊沉积物中磷的周转时间都较短,并且湖中往往出现养分的“短路代谢”,所以通常生产力较高。根据水体中物质循环的规律,努力使系统中的养分得到充分的循环和利用,是提高渔业水体生产力的一项重要措施。中国正在推广的桑基鱼塘,用鱼塘底泥作为桑田肥料,桑叶养蚕,蚕砂(粪)用于施肥养鱼。这在调节营养物的循环过程,充分利用自然资源和维护自然生态平衡方面,已取得良好的效果。(见农业生态)
能流
按国际生物学规划(IBP)的广泛调查结果,世界湖泊的总初级生产力变动在500千卡/(米2·年) (北极湖)和1万千卡/(米2·年) (某些热带湖泊)之间。河流生态系统的初级生产力较低,除某些泉流外,通常很少超过1000千卡/(米2·年)。据估计,世界湖泊与河流的总净初级生产量为每年0.6×1016千卡,占生物圈总净初级生产量的0.82%,平均净初级生产力为每年每平方米2250千卡。
淡水生态系统中的能流通过牧食食物链、捕食食物链和碎屑食物链共同实现。但在不同的水体中,这两类能流途径所起的作用往往有明显差别。大型湖泊和水库中以牧食食物链为主,而碎屑食物链在水生高等植物繁茂的水体中起主导作用。在后一类水体中,90%的初级生产量通过碎屑路线被利用。水体中的碎屑,除了来自死亡的动植物体之外,植食性动物的排粪是另一个重要来源。
食物链原理对淡水渔业生产有重要的指导意义。渔业生产的基本目标,是充分利用水体的饵料生物资源,最大限度地提高鱼产量。为此,应采取一系列措施来调节水体的生产过程,使系统中的能源物质尽可能转化为鱼产品。这些措施包括:
(1)多放养食物链短的鱼类,以有效地利用生产者所固定的能量;
(2)控制凶猛鱼类种群的发展,以减少能量多环节传递所造成的损失和对放养鱼类的危害;
(3)注意天然杂食性鱼类的繁殖保护,并适当增加食碎屑鱼类的放养量,以提高水体中碎屑资源的利用率。目前,以鲢、鳙为主体的多品种放养,已成为中国淡水渔业生产上成功的经营方式。
演替与平衡
生态系统的发育,即通常所讲的生态演替,指一定区域内连续进行的前一群落被后一群落所替代的发展过程。演替起因于系统内生物导致的环境改变、外来种的入侵等生物学过程,或起因于气候变迁、土壤侵蚀、河流泛滥、环境污染等外部力量,也可能是这两方面因素共同作用的结果。演替过程中依次出现的一系列群落构成一个演替系列,其中早期出现并且存在时间比较短暂的群落属于先锋阶段,相继出现的群落属于发育阶段或演替阶段,而最终形成的稳定系统称为演替顶极。演替是群落发育的有序过程,这一过程基本上有一定的规律,因此通常可以预测。
淡水生态系统发育的基本模式,是从贫营养到富营养和由水体到陆地。这一过程通常可用湖泊的发育史来说明。湖泊形成初期,养分含量很低,常常仅有一些浮游生物。由于外源物质的输入,湖中有机物质含量增高,并出现沿岸沉积物。于是,水生高等植物慢慢发展起来,水生昆虫、环节动物、鱼类等多种动物陆续迁入。随着时间的推移,水底沉积物不断增加,沿岸带植物逐渐向湖心发展。因此,湖泊逐渐由深变浅,由大变小,直至整个水体最后完全消失。淡水生态系统的发育过程一般比陆地生态系统缓慢,并且往往因外源物质的大量输入而出现十分复杂的情况。同时,在发育过程中,生态系统内一些较小的群落单元,如浮游生物群落、微生物群落等,常常表现明显的季节演替现象。
生态系统是天然的控制系统,其自我调节依赖于负反馈机制。在淡水生态系统中,许多反馈路线存在于生物与生物和生物与环境之间,通过反馈,种群大小得以维持在一定的限度内。例如,当某种凶猛鱼或浮游植物种群增长得过大,以致其食物或养分供应不能满足时,就会出现大量的自然死亡,种群繁殖率下降,以及凶猛鱼残害其幼体的现象,结果种群数量倾向于减少。除捕食和竞争外,生态系统中所发生的其他相互影响,如寄生、共生等,都有维持系统相对稳定的趋向。
然而,生态系统的自我调节能力是有限度的,即使是处于成熟阶段(即演替顶极)的生态系统,也只能承受一定强度的外来干扰(压力)。当外部压力超过所允许的限度时,就会破坏生态系统的平衡状态,甚至造成整个系统的崩溃。工业污染物倾泻入湖造成的湖泊生物大量死亡和湖泊生态系统的破坏(见污染生态)以及外源养分的输入过多所引起的湖泊富营养化,都是自然生态平衡遭到破坏的典型实例。
大量的外源养分(磷、氮等)进入湖泊后,加速了藻类(主要是蓝藻)的生长,水体中的有机物质大幅度增加。可是,蓝藻不能很好地被植食性动物所利用,以致水体中出现初级生产的相对过剩。同时,大量的有机物质沉积于水底,其分解造成湖下层严重缺氧。结果,很多水生动物逐渐消失,而藻类生物量越来越大,水质不断恶化。在富营养化水体中,由于群落结构的破坏和一系列功能失调,物质循环和能流过程不能正常完成。
外源物质人为地大量输入所引起的水体富营养化,与天然情况下的水体富营养化根本不同。后者是在系统内生物学过程起主导作用的情况下产生,其变化过程是渐进的,漫长的,反映了生态演替的正常趋向。而人为的水体富营养化,则是在外部力量的影响下产生的;由于系统内的生物学过程受到严重干扰,因而生态演替的正常趋向发生逆转。在人为的富营养化水体中,物种的多样性明显下降,采 r型策略的物种重新占优势,这可看作典型的演替逆转现象。
20世纪70年代以来,由于人类经济活动的影响,大量的淡水水体出现污染或富营养化,致使人类的生活和生产用水受到严重威胁,淡水生物资源遭到极大的破坏。因此,加强对淡水生态系统的保护,采取有效措施防治水体污染或富营养化,已成为关系到人类的生存和社会经济发展的一个重大问题。在水体富营养化的治理中,通过采取污水截流、换水、挖泥等措施,已在不同程度上收到了改善水质的效果。实践证明,只要减少或停止外源养分的输入,就可以大大延缓或终止水体富营养化的进程。
- 参考书目
- R.K.Barnes, K.H. Mann,Fundamentals of Aquatic Ecosystems, Blackwell Scientific Publications, Oxford,1980.
- E.P.Odum, Fundamentals of Ecology, 3rd ed.,W.B.Saunders Co.,Philadelphia,1976.