[拼音]:liansuo he jiaohuan
[外文]:linkage and crossing-over
位于同一染色体上的基因伴同遗传的现象称为连锁。由于同源染色体相互之间发生交换而使原来在同一染色体上的基因不再伴同遗传的现象称为交换,在原核生物中多称为重组。连锁和交换是生物界的普遍现象。
连锁对于生命的延续是十分必要的。因为一个细胞中有许多基因,如果它们各个分散,便很难设想在细胞分裂过程中怎样可以使每一个子细胞都准确地获得每一个基因。交换对于生物的进化有重要意义,它可以使配子中的基因组合变化无穷,从而带来生物个体间的更多的变化,为自然选择提供更大的可能性。
交换一般是对等的,可是也有不对等的。不对等交换导致少量染色体重复,这被认为是生物进化中新基因的主要来源之一。此外,交换和育种工作也有密切的关系。重组机制的研究是遗传学中的一个古老的问题,现在已经成为分子遗传学研究中的吸引人的课题。
物质基础1906年英国遗传学家W.贝特森等在香豌豆的杂交试验中发现紫花基因(purple,P)、红花基因(p)、长花粉基因(long,L)和圆花粉基因(l)在子二代中的分离比都各自符合于3:1(见孟德尔定律),可是两对基因的分离比却不符合于 9:3:3:1。似乎两对基因在杂交子代中的组合并不是随机的,而是原来属于一个亲本的两个基因更倾向于出现在同一配子中。1911年美国遗传学家t.h.摩尔根根据在果蝇中发现的类似现象提出连锁和交换概念,认为倾向于伴同遗传的基因位于同一染色体上,而因同源染色体间的交换又使这些基因不再伴同遗传。
1913年美国遗传学家A.H.斯特蒂文特首先在果蝇中测定了六个基因在X染色体上的排列顺序,获得了第一个连锁群,奠立了遗传的染色体学说。根据这个学说,可以预期连锁群数等于单倍染色体数(见染色体倍性)。在深入研究过的生物中,都证实了这一假设。例如人、小鼠、黑腹果蝇的染色体数分别是(46,XY)、(38,XY)和(8,XY),它们的连锁群数都和单倍染色体数相符,分别是23、19和4。玉米、水稻、豌豆的染色体数分别是20、24和14,它们的连锁群数分别是10、12和7;大肠杆菌是单倍体生物,只有一个环状的染色体,它的连锁群数是1。
根据这一学说,还应该在显微镜下看到和交换有关的染色体图象。1909年F.A.詹森斯首先在两栖类动物和直翅目昆虫生殖母细胞的减数分裂中,看到了二价体中的交叉,并提出交叉型学说,认为交叉是交换的结果而不是交换发生的原因。也就是说遗传学上的交换发生在细胞学上的交叉出现以前,而并不是由于染色单体相重叠而导致交换的发生。按照交叉型学说,交换在粗线期甚或更早时期已经发生;在双线期中两个未分裂的着丝粒分别带着两个姊妹染色单体趋向两极时使已经发生交换的位置上出现交叉。按照经典学说,则交换将发生在双线期中或更早时期的二价体上两个染色单体相重叠的部位,所以按照不同的学说,同一个二价体图像中的四个染色单体间存在着不同的关系。交叉型学说已经得到公认。
在光学显微镜下,交叉首先在双线期中看到,可是在电子显微镜下在偶线期-粗线期就可以看到称为联会线复合体的结构。它的显微图像在各种生物中不完全相同,但是基本上包括两条侧线和一条中轴,中轴和侧线之间往往可以看到许多横纹。许多事实都说明这一结构和联会与交叉有关。例如酵母菌的细胞分裂周期突变型(celldivision cycle,cdc)cdc7-1和cdc7-4,在较高的限制性温度中既不发生交换,也不形成联会线复合体;把这些突变型放到较低的非限制性温度中则发生交换,同时出现联会线复合体。隐性无交叉基因 C(3)G为纯合型的雌性果蝇中不发生交换,同时在它的减数分裂中看不到联会线复合体。不过联会线复合体的存在只是交换的必要条件而不是充分条件,因为在番茄和大麦的一些无交叉突变型细胞中可以看到联会线复合体,可是却不发生交换;在雌性家蚕卵母细胞的减数分裂中可以看到联会线复合体,可是同样不发生交换。
发生的位置一般情况下染色体愈长则显微镜下可以看到的交叉数愈多。但交叉数的多少又不完全决定于染色体的长度。在某些生物中,在某一长度范围内染色体长度和交叉数呈正比;如果低于这一长度范围,则实际交叉数比预期数大。例如根据染色体长度看来预期交叉数不到一个的二价体上都出现了一个交叉。在红腿蝗(Melanoplus femurrubrum)、风信子(Hyacinthus ame-thystinus)、丝兰(Yucca flaccida)等生物中都报道过这一现象。这说明任何一个染色体不论它是怎样短,都必然发生一个交叉。这种机制现在还不清楚,一般认为它对于细胞分裂过程中染色体的有规律的行为起着重要的作用。
在某些生物中,交叉在染色体上的分布倾向于集中在某些部位。例如大麦、燕麦、鸭跖草、月见草等的交叉倾向于发生在染色体的两端;贝母属(Fritillaria)植物的交叉较多地发生在着丝粒的两旁;黑腹果蝇(Drosophilamelanogaster)则恰好相反。在黑腹果蝇的细胞学图上基因的分布是均一的,可是在遗传学图上则比较多的基因位于着丝粒两旁。对于这一现象的一个合理的解释是交换较少发生在着丝粒的两旁。
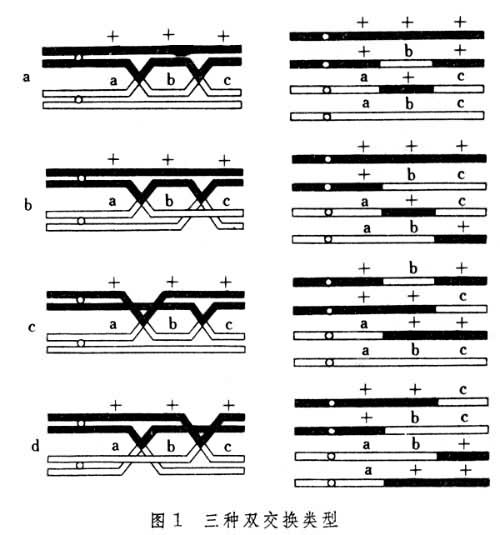
同一染色体如果出现两个交叉,就交换所涉及的染色单体来讲,邻接的两个交换的关系有三种:包括两条染色单体的两线双交换(图1a),包括三条染色单体的三线双交换(图1b、c),包括四条染色单体的四线双交换(图1d);它们分别带来1%、50%、100%基因重组;四分体类型分别属于包括两种类型的亲代二型(PD)、包括四种类型的四型(T)和包括两种类型的非亲代二型(NPD)。
1916年美国遗传学家h.j.马勒在果蝇中发现干涉现象。一个位置上的一个交叉对于邻近位置上的交叉发生的影响称为干涉,如果一个交叉的发生减少另一交叉发生的概率则称为正干涉,如果增加后者发生的概率则称为负干涉。前一交叉影响邻近位置上另一任何形式的交叉的现象称为染色体干涉(交叉干涉);前一交叉所涉及的两条染色单体影响另一位置上相同的两条染色单体再次发生交换的现象称为染色单体干涉。
正交叉干涉的结果使实际发生的双交换频率低于假定两个交换的发生互不干涉所预期的频率。例如实际观察到基因a和b之间的交换频率是X,基因b和c之间的交换频率是 Y。那么同时发生在这两个位置的双交换的预期频率应是XY。如果实际双交换频率低于预期值,说明有正交叉干涉。实际双交换频率与预期双交换频率之比称为并发系数。如果实际数值和理论值相同则并发系数是1,说明不存在交叉干涉。对粗糙脉孢菌进行遗传学分析的结果说明不同染色体臂之间没有正交叉干涉,同一染色体臂上则存在正交叉干涉,而且基因距离愈近干涉程度愈大。正交叉干涉的机制目前还不清楚。
负交叉干涉现象曾在家蚕和λ噬菌体等中发现。在λ噬菌体中,发现双交换的并发系数往往大于5,说明一个位置上发生了一次交换以后,它的附近位置上更容易发生另一次交换。已经提出一些模型试图说明负交叉干涉的机制,但都还需要进一步验证。
如果不存在染色单体干涉,那么两线双交换、三线双交换和四线双交换频率之比应是 1:2:1。染色单体干涉的结果使两线双交换减少,从而使子囊菌杂交子代中亲代二型子囊减少,或者使任何生物的杂交子代中重组型子代超过50%,这一现象曾在酵母菌和另一些真菌中发现。
发生的时间在细胞减数分裂的过程中同源的两条染色体发生联会,然后各自分裂,形成包括四个染色单体的二价体。在交叉位置上可以看到只有两条染色单体发生了交换(图1),这是说明交换发生在四线期的细胞学证据。遗传学证据来自四分体分析。如果交换发生在染色体分裂以前,那么通过减数分裂所产生的四分体将是非亲代二型(NDP),不发生交换的四分体将是亲代二型(PD),但是不可能出现四型(T)。只有交换发生在染色体已经分裂,即发生在四线期才有可能得到四型四分体。在高等动植物中,减数分裂的产物是混杂的,很难判断交换发生的时间,在子囊菌中可以得到三种四分体,因此说明交换发生在四线期。
关于交换发生的机制曾经有两种学说。一种学说称为样板选择,于1931年由美国细胞遗传学家J.贝林提出,认为交换是染色单体复制过程中变换模版的结果。另一种学说称为断裂-重接,于1937年由英国细胞学家C.D.达林顿提出,认为交换是复制完成后两个染色单体断裂并重新连接的结果。至少在噬菌体中,已经有实验证明交换可以由于断裂重接发生。
影响因素 环境因素许多环境因素影响交换,包括温度、水分、营养、射线、化学药品等。早在1917年已经报道了温度对于果蝇的交换的影响,发现22℃中饲养的雌性果蝇的交换频率最低,温度低于22℃或高于22℃时交换频率都显著提高。
基因型在果蝇中很早便发现性别对于交换的影响,交换只发生在雌蝇而不发生在雄蝇中。家蚕则恰恰相反,交换只发生在雄蚕而不发生在雌蚕中。实际上凡是性别为性染色体所控制的生物,异配性别个体中一般总是较少发生交换。早在1922年英国生理学家兼遗传学家j.b.s.霍尔丹提出了一条定律:凡是较少发生交换的个体必定是异配性别个体。这一定律称为霍尔丹定律。同一年在果蝇中报道了隐性交换缺陷型C(3)G。抑制交换或者促进交换的突变型在动物如果蝇和人,在高等植物如小麦和玉米,在真菌如脉孢菌、曲霉、酵母菌等以及在λ噬菌体等中都有报道。
在果蝇和黑麦中发现的影响交换的突变型可以大致分为两类:一类突变型显著地减少交换,可是不影响染色体上交换位置的分布;另一类突变型改变交换的分布,可是不一定减少交换。后一类突变型可以理解为只是影响导致交换的必要条件(如影响染色体的联会等),而前一类突变型可以理解为直接影响交换的过程。
许多影响交换的突变型往往同时是切除修复缺陷型,对于紫外线的杀菌作用格外敏感;某些突变型是重组修复缺陷型;另一些突变型如果蝇的C(3)G则极易为X射线诱发而产生新的突变。这些事实说明交换和 dna损伤修复过程有某些共同的地方。
原核生物的染色体和真核生物的染色体有一些本质上的区别,但同样发现了一些影响交换的突变型,而且也常是对紫外线敏感的;例如大肠杆菌重组缺陷型(recom-bination deficient,rec A)等。
交换和基因重组如果基因在染色体上作线性排列,那么染色体交换应该带来基因重组。基因重组和染色体交换之间的平行关系的最直观的证据来自玉米和果蝇的细胞遗传学分析。玉米的每一个染色体都可以辨认。在某一个玉米品系中,第9染色体的一端有一个显著的染色节,在另一个黄绿色(yellow green,yg)、蜡质(waxy,wx)品系的同一染色体的另一端有较长的一段易位染色体。基因yg和wx位置在第9染色体的有染色节的臂上。在杂交子代中可以一方面观察遗传性状之间的组合,另一方面观察染色体形态的改变。1931年H.B.克赖顿和美国遗传学家B.麦克林托克观察到凡是基因发生了重组的子代,它的染色体的形态也不同于双亲。这些结果只能用基因重组来源于同源染色体间的交换来说明。同一年美国动物学家兼遗传学家C.斯特恩在果蝇中得到同一结论。
由于交换发生在四线期,所以一次交换只导致半数染色单体上的基因发生重组。两线双交换并不带来重组,四线双交换带来全部染色单体的重组,三线双交换则带来50%重组,所以和一次交换一样,两次交换同样导致半数染色单体上的基因发生重组。三次、四次或更多次交换也只导致50%重组。如果两个基因不在同一染色体上,那么它们之间的组合必定是随机的,也就是说一个双基因杂合体的减数分裂产物中,亲本类型和重组类型各占50%。但是两个在同一染色体上的基因不论距离是多么远,重组值以50%为极限。所以重组值显然小于50%是基因连锁的证据,重组值是50%则并不是基因不连锁的证据。但是在两个相距较远的基因中间必然可以找到一些基因,它们和这两个基因的重组值将小于50%。整个连锁群就可以这样逐步建立。
自然群体中的交换和重组每一种生物细胞中的染色体数是固定的,每一个细胞中全部染色体的交叉的平均数也是比较恒定的。异花授粉的黑麦(Secalecereale)经人工自交可以得到交叉平均值低于亲本的一些自交系。这些事实说明交换的发生为基因所控制,交叉数的遗传控制是多基因的。
在交叉平均数较低的黑麦中交叉常限于染色体的两端,在交叉平均数较高的黑麦中额外的交叉位置在染色体的中部。通过自交也可以得到交叉的平均数虽少,可是多数交叉位置在染色体中部的品系。这又说明交叉的位置也由基因所控制,这控制也是多基因的。
比较多花毒麦(Lolium multiflorum)的各个自然群体的交叉平均值和开花时间的变化程度,发现变化愈小者交叉平均数愈大。这是因为交叉平均数愈大则交换发生得愈多,由基因重组带来的变异也愈多,在长期的自然选择下极端类型多被淘汰,所以留下的个体中变化程度较小。在草地羊茅 (Festuca pratensis)和多年生毒麦 (Loliumperenne)中也有类似的报道。这些事例都能说明交叉平均数大的群体的长期可塑性大,可是短期适应性较差;平均数小的群体则恰好相反。
交换除了受到基因的控制以外也和染色体结构有关。在果蝇中已经证明倒位杂合体中倒位能有效地抑制倒位区域内的交换(见染色体畸变)。在许多果蝇群体中保持着这种染色体杂合状态,说明这种基因组合在特定生态条件下有利于生存(见遗传多态性)。
染色体相互易位同样可以起到保持某种基因组合而减少重组的作用(见染色体畸变)。在加州芍药(Paeoniacalifornicae)和美洲蜚蠊 (Periplaneta americana)中都曾发现某些自然群体中保持高度的易位杂合状态。一个极端的例子是一些拉马克月见草 (Oenothera la-marckiana)的染色体易位杂合体,这里易位所涉及的染色体是7对,在它们的减数分裂中期可以看到多达14个染色体形成的环,它们在中期的排列有90%属于交替式,并且交换都发生在染色体的两端。非但如此,它们的纯合体往往是致死的。这样一种平衡致死(见致死突变型)体系进一步保证杂合状态。并且实质上使所有的染色体成为一个“连锁群”,而很少可能通过交换而在“连锁群”间发生基因重组。
各种形式的交换和重组体细胞交换于1936年首先由C.斯特恩在黑腹果蝇中发现。黄体(yellow body,y)和焦刚毛(singed,sn)这两个X连锁基因都呈杂合状态的雌性果蝇+y/sn+的表型是野生型。可是在少数果蝇身上可以观察到孪生斑,即两小块邻接的组织:一块上显示突变型黄体性状,但上面的刚毛是正常的;旁边一块组织上的刚毛弯曲,但体色却是正常的。这一现象不能用基因突变,而只能用体细胞交换来解释(图2)。

构巢曲霉是一种产生有性孢子的子囊菌,它的营养体是单倍体。两个单倍体的菌丝联接以后产生异核体。在异核体菌丝体中的个别细胞核融合成为二倍体核,由二倍体分生孢子萌发产生二倍体菌丝体。如果这一个二倍体菌株来自两个分别具有不同基因标记的单倍体菌株,那么就可以看到它的菌落上会出现通过体细胞交换而产生的扇形变异部分。这一全部过程称为准性生殖过程。准性生殖过程中所发生的体细胞交换,可以用来进行基因定位和杂交育种。
不等交换交换一般是对等的,即两个同源染色体在对应的位置上发生交换,发生了交换的两个染色体都不增加或减少任何基因。如果同源染色体联会时不是准确地配对,那么交换发生在不对应的位置上,结果所得到的两个染色体中就有一个染色体缺少了一小部分,而另一染色体多了一小部分。这种交换称不等交换(见位置效应)。通过不等交换所出现的这种多余的或重复的基因如果发生突变也不会影响细胞的生存,因为原有的基因可以执行正常的功能。这样,新的基因便可以通过多余基因的一再突变而形成。因此,不等交换被认为是进化过程中新基因的主要来源之一。
细菌的重组大肠杆菌和枯草杆菌都只有一个呈环状的染色体,即一个连锁群。在细菌染色体的重组过程中既不出现交叉,也没有联会线复合体,也不通过减数分裂而产生四分体。而且交换只发生在以单链形式进入受体的脱氧核糖核酸 (DNA)片段中。尽管重组机制有所不同,但是真核生物和细菌染色体的重组都是以整段染色体的联会作为前提,这种重组方式称为同源重组或一般重组。
非同源重组,是发生重组的两个部分中,它们只有极小一部分同源结构。它们在发生重组以前,不像减数分裂中的同源染色体那样整段染色体进行联会,而是联会仅发生在少到十几对核苷酸之间,而且重组的结果是一段DNA插入到另一DNA分子中。
λ噬菌体整合到大肠杆菌染色体上的过程便是一种非同源重组(见转导),它只整合在半乳糖基因 (galac-tose,gal)和生物素基因(biotin,bio)之间,所以又称为位点专一重组。转座因子的整合也属于非同源重组,可是它们的位置专一程度不一。大肠杆菌中的插入顺序IS4是绝对专一的,Mu-1噬菌体则是绝对不专一的,多数转座子的专一性介于两者之间。
λ噬菌体的整合过程中并不进行复制,所以这种重组又称为保守性重组;转座子的整合过程包括它本身的复制,所以这种重组又称为复制式重组(见转座因子)。
姊妹染色体单体互换指来自两个亲本的染色体各自分裂成为两个姊妹染色单体后发生在姊妹染色单体之间的交换。早期的许多遗传学和细胞遗传学研究结果说明姊妹染色单体之间不发生交换。J.H.泰勒于1958年应用同位素标记方法,在Bellavalia细胞的有丝分裂过程中发现有姊妹染色单体互换现象。以后应用5-溴尿嘧啶脱氧核苷(BrdU)标记后再经荧光染料或吉姆萨染料染色方法在许多生物中证实了这一现象(见dna损伤修复、毒理遗传学)。不过,至今还没有确切的证据足以证明姊妹染色单体互换在通过减数分裂发生的基因重组方面起着任何作用,它可能只是和DNA损伤有关。
实践和理论意义 育种育种工作中的一个课题是取得种内杂交亲本的优良基因组合。如果要求组合在一起的两个基因的座位是紧密连锁的,那么在杂交子一代的减数分裂中通过交换而出现所要求的基因组合的配子将是稀少的。这就需要从为数较多的子代中进行选择才可能得到所要求的基因组合。根据已知的连锁关系,可以预测在多大的子代群体中才能发现所要求的基因组合,从而可以减少育种工作中的盲目性。作物育种工作的另一个课题是通过远缘杂交从野生植物中引进耐干旱、抗虫、抗病等基因。要引进这些基因就必须使这些植物的染色体和栽培植物的染色体发生交换。可是由于长期进化的结果,即使某一栽培植物和某一野生植物来自同一祖先,它们的染色体也不过是部分同源的,因而不易发生交换。因此必须研究如何促进部分同源染色体的交换才能取得预期的效果。小麦的ph基因阻碍小麦-冰草杂种减数分裂中部分同源染色体的联会。设法使ph基因缺失或受到抑制,就能使部分同源染色体间发生交换。通过这一方法,至少已经把三个冰草的抗病基因引进到小麦中去。
产前诊断一个妇女从她的父亲得到一个带有血友病基因(hemophilia,hm)和葡萄糖6-磷酸脱氢酶(G-6PD)座位的同工酶A基因,由于已经知道这两个基因的距离是五个图距单位,如果通过羊水检查而发现同工酶A的存在,并且由核型分析知道胎儿是男性的,就可以预测这一胎儿患有血友病的机率是95%,因为G-6PD基因和hm基因之间发生交换的机率是5%。相反,如果羊水检查中不能查出同工酶 A的存在,那么这一胎儿患有血友病的机率是5%,因为不带有同工酶A基因的X染色体中,只有5%带有血友病基因。
理论意义鉴于交换和重组在遗传学及生产实践中的意义,长期以来是遗传学研究中的一个中心课题。近年来交换的分子机制的研究特别受到注意。在这方面噬菌体的简单的体制和子囊菌的一次减数分裂所产生的四分体,以一定顺序排列在子囊中等特点在交换的研究中尤其受到重视并被利用。在λ噬菌体整合到大肠杆菌染色体上的过程中,发生交换的部位称为噬菌体附着位点(attP)和细菌附着位点(attB)。两个附着位点完全同源,并且实际上发生交换的部位只有15个核苷酸对。这一简单的体制适合于进行重组过程的分子遗传学研究。子囊菌和其他生物中影响减数分裂的突变型也被广泛用来研究交换过程。已经发现一些影响交换的突变型和 dna损伤修复有关,而DNA损伤修复又和基因突变有关,甚或与癌变和衰老有关。所以对交换分子机制及其他基本生命活动的关系的研究,都将是今后的重要课题。
- 参考书目
- H.里斯等著,张勋令译:《染色体遗传学》,科学出版社,北京,1983。(H.Rees,R.N.Gones,Chromosome Genetics,Edward Arnold Ltd.,New York,London,1977.)