[拼音]:shengwu fenbu
[外文]:distribution of organisms
即生物的地理分布。任何生物分类单位(种、属、科或种以下的亚种、变种等)在地球表面分布的范围叫做它的分布区,不同分类单位的分布区大小和形状差别很大,这既取决于生物本身的散布能力,也取决于它所遇到的外界条件;此外,还决定于历史因素等。
生物的散布能力是由它的繁殖速度和它的散布方式决定的。凡产生种子或孢子多、繁殖快的,它的散布能力就大,低等植物往往产生惊人数量的繁殖体,它们的散布能力就比产生较少繁殖体的种子植物大得多。动物也是一样,每窝仔仅数个的哺乳动物(灵长目、鲸目等),其散布能力远远不如大量产卵的昆虫。风布和动物布与不利用外界能源的散布(重力散布、营养繁殖)方式相比,能到达的距离更远。对于少数生物,水布也是远距离散布的一种方式。在交通发达的现代,人类帮助某些生物的散布,其速度之快,所达距离之远,超过其他动力。
限制生物分布的因素
生物的本性是产生尽可能多的后代,占领尽可能大的地盘(分布区),然而由于外界条件的限制,它并不能实现这一本性。限制生物分布的外界因素来自两方面:环境影响;同其他生物的相互作用。
环境影响生物只能生存于一定的环境条件范围内。一般说来,气候和土壤是决定植物分布的主要环境条件;大多数动物以植物为食或以其为蔽所,因此动物的分布往往与植物的分布相合。从赤道到极地和从沿海到内陆的气候变化不仅伴随着植物分布的改变,而且也伴随着动物分布的改变。
有时分布区的外形不决定于气候,而决定于基质。例如盐生植物的分布与盐渍土的分布相一致。
环境影响有时表现为明显的“地理障碍”。例如,海洋对于绝大多数陆生生物是不可逾越的障碍,陆地对于水生生物也是一样,所以许多生物的分布区以岸线为界。苏伊士运河未凿通前,地中海和红海具有极不相同的动物区系,运河挖通后,某些红海种的分布区延伸到地中海东部。
地形对于陆地栖居者来说也是重要的分布障碍,连续不断的山链(例如喜马拉雅山、秦岭、天山、祁连山)对于生活在低海拔地区的种有如一堵不可通过的墙,因此山两边可以发现很多不同的种类。大型湖泊和河流有时也成为陆地栖居者通过的障碍。
同其他生物的相互作用一个生物种的分布可能决定于同其他种的相互作用。例如有毒植物牛心朴子目前在鄂尔多斯一带广泛分布,这不仅由于它本身不为牲畜所吃,还由于该地区长期强度放牧,减少了其他植物对它的竞争压力;各类针茅数量的日渐减少则是牲畜直接大量啃食所造成的。
一定植物群落常常成为某些动物分布的限制,特别是占领广阔空间的群落,这种限制作用尤其显著。例如,对于大多数热带森林动物来说,稀树草原是它们分布的障碍,但同时热带森林也是稀树草原栖居者分布的障碍。在这种情况下,障碍作用既是环境条件的改变,也是竞争的结果。
生物的局部分布
从小范围看,生物个体分布在不同种之间差别很大,一般有3种基本类型(图1):
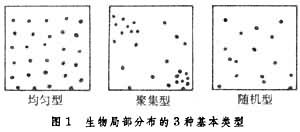 均匀分布
均匀分布
个体间大致等距离分布,一些占域鸟和森林中的树木表现为这种类型。中国北疆古尔班通古特沙漠中的白梭梭和美国索诺兰沙漠中的巨仙人柱是这种分布的最典型代表。均匀型的产生往往是由于个体相互排斥,因而彼此保持一定距离。
聚集分布个体成群或聚集分布。聚集型形成的原因有几种:或者由于个体相互接近有利(如动物群);或者由于环境的不均匀性,有利于生物在某一部分存在而不利于在另一部分存在;或者由于繁殖的特性造成(植物的匍匐茎或根茎的无性繁殖)。大多数动物趋向于聚集型分布。土壤或森林枯枝落叶层中大多数节肢动物都成群或聚集分布。
随机分布个体的分布是偶然性的。在特别一致的环境里,生物自由选择定居地点,这有利于随机型分布的出现。例如在潮带间的泥滩,某些蛤类通常呈随机型分布。
生物的大范围分布
由于起源、历史、生态要求和环境条件的差异,不同物种分布区的大小和外形极其多种多样,但几乎所有物种都在地球表面占据不规则形状的面积。分布区形状的不规则性在大多数情况下由环境特点所决定。
世界性分布区指覆盖地球大部分面积的分布区。目和科的世界性分布较多。动物中的轮虫类,种子植物中的菊科、禾本科、豆科和莎草科都具有世界性分布。属和种只有很少是相对世界性的。大约有20种左右种子植物可以归入世界性分布,它们包括许多水生和沼泽植物,如浮萍、芦苇、香蒲、水烛等。伴随人类散布的杂草,有些也是世界性的,如早熟禾、荠菜、蒲公英、大车前等。动物中的家蝇和褐家鼠等的分布也遍及全世界。
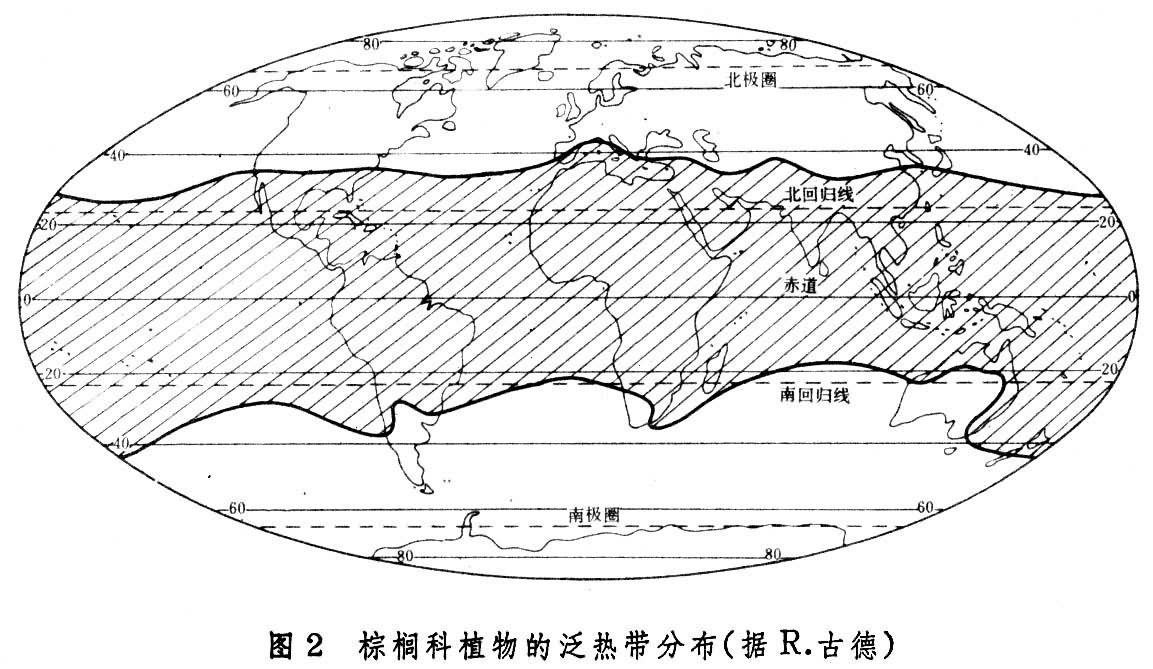
环大陆和环海洋分布区指占据某一纬度范围的陆地或海洋的分布区。包括环北极分布,如拉伯兰苔草;北半球中纬环陆分布,如驴蹄草,茶藨子;泛热带分布,如棕榈科植物(图2)、含羞草、马钱子、羊蹄甲等;南半球高纬环陆分布,这是断续分布,以某些古老类群的生物为特征,如原始导管植物,有爪纲以及Styloniceae科的等足目甲壳类。
 特有分布区
特有分布区
与世界性分布相反,特有分布只限分布于某一有限地域。特有分布区的面积可能多种多样,目、科、属的特有分布区面积一般较大,例如澳大利亚、塔斯马尼亚和新几内亚总起来形成单孔目的特有地域。对于种或变种来说,其特有分布区可能面积很小。例如就现在所知,金佛山兰(Tangtsinia nanchuanica)只产于中国四川省南川县的金佛山。
从进化观点看,特有现象可能是两种完全不同的进化过程的结果:
(1)古特有种,或称残遗特有种,都是古老的物种,早先的分布区极为广阔,后来由于气候改变或由于适应力较强的其他种的竞争,其分布区逐渐缩小。所以古特有种大多处于消失的过程中。例如银杏,在中生代三叠纪和侏罗纪时遍布全世界,在第四纪冰川期,中欧和北美的银杏都已绝灭,只在中国浙江天目山一带还保存有野生种。所以银杏目前的分布区(野生)很小,是古特有分布区。水杉(Metasequoia glyptostroboides)在第三纪时也广布于北半球,现在野生状态的个体仅在中国湖北西部利川县和重庆万州的少数几个地区能够见到,也是古特有种。被隔离的山链往往是特有种集中的地方。例如在非洲撒哈拉山区的阿哈加尔山脉,特有种占植物的40%。
(2)新特有种,或称年幼特有种,是一类不久以前在局部地区产生的特有种。由于形成时间不久,往往只达到亚种级。例如生活于不列颠和中欧湖泊中的一种淡水鲑鱼鸦巴沙(Coregonus),在每一湖泊中都有不同形态的个体。在不久以前才与欧洲大陆分离的不列颠群岛上,陆生动物中新特有种数目特别多。
替代分布区从相邻两个区域的同样生境起源,或从同一区域的两个不同生境起源的两个近缘分类单位相互取代的现象叫替代现象,它们的分布区是替代分布区。前一种情况是地理替代;后一种情况是生态替代。前者如地中海流域的油橄榄(Olea europaea),它的野生形式在撒哈拉山区为非常接近的O.laperrinei所替代;后者如中国南方的狗脊 (Woodwardia japonica)和镰狗脊W.unigemmata),狗脊生长于酸性土上,而镰狗脊则要求石灰性土。在砂质岩(或第四纪红层)和石灰岩相间分布的山坡,就可以看到狗脊和镰狗脊相间出现。巴拿马地峡东岸和西岸动物区系的相互替代是海洋替代的极好例子。地峡大约在上新世开始形成,使两岸相互隔开。目前许多鱼和软体动物可以相近形式成对出现,其中每一对中的一个出现在大西洋近岸水域,另一个就出现在太平洋近岸水域。
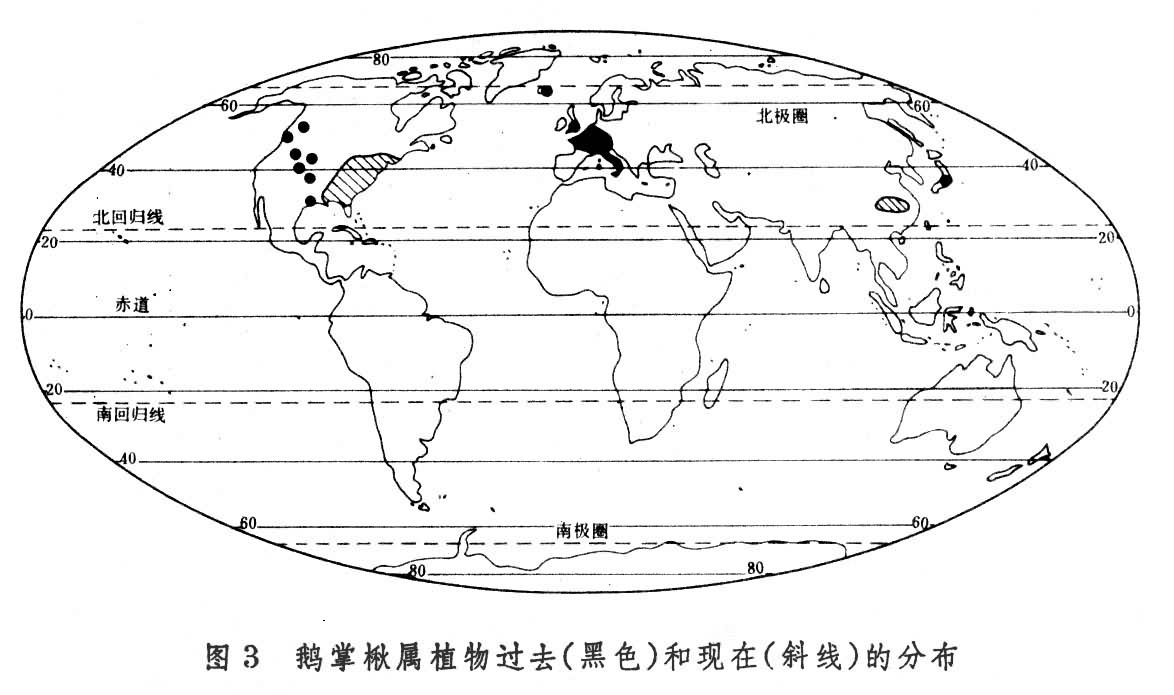
间断分布区或称不连续分布区。有些分类单位的分布区断裂成几个部分,并且彼此相距很远。相似的或有密切关系的甚至相同的种出现在地球的不同部分,形成相互隔离的分布范围,这样的分布区叫间断分布区或不连续分布区。间断分布区有多种多样的类型。例如,著名的第三纪残遗植物鹅掌楸属中,一种(即鹅掌楸)分布于中国南方,另一种(北美鹅掌楸)分布于北美东部,为洲际间断(图3)。不仅鹅掌楸属,整个木兰族也是洲际间断。
许多种具有北极-高山间断分布,即它们的主要分布区在北极附近的冻原,但也出现在中纬度的高山上。例如对生叶虎耳草,既见于北极地区的低地,也出现在欧洲南部高山的高海拔生境。许多北极性的植物如高山唐松草(Thalictrum alpinum)、 珠芽蓼 (Polygonum vivipa-rum)、虎耳草(Saxifraga hirculus)、萝蒂草(Loydiaserotina)等在中国西南高山也能见到。
特别令人惊奇的是两极间断分布(或称双极分布)。同一个属的不同种,甚至同种,既出现在北极附近,也出现在南极附近,而中间地区却没有,例如岩高兰属主要具有环北极分布区,但又出现在南美的南端。海洋动物中也有双极分布的例子,例如北极海参(Psolus)和南极海参。
关于造成间断分布的原因曾有许多争论。一种解释是远距离散布,特别是往返于南北半球之间的候鸟的迁徙,可能是传布繁殖体的重要媒介。候鸟迁徙对解释某些植物的两极间断分布有帮助,但不能解释有些洲际间断,如上面谈到的鹅掌楸在东亚和北美东部之间的间断。为什么同样纬度、气候条件相似的北美西部没有鹅掌楸,这难以用候鸟迁徙解释。有些人提出相似种在地表不同的和远离的部分完全独立地起源的假说,但很少证据能够证明这点。普遍接受的假说是:相似的或遗传上有关系的生物类型在过去必然来源于一共同祖先,从共同的起源中心向外侵移到它们所占据的地区,而目前分布区的不连续性是以后发生的环境改变──“障碍”造成的,这种障碍把它们目前占领的地区分开。例如,北极-高山间断分布是由于在冰川期间北半球的冰盖向南延伸到中纬度,覆盖了欧亚大陆和北美的广大地区(图4)。
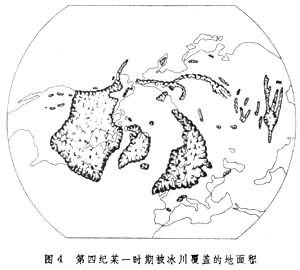
随着冰川的南侵,原来分布在北极附近的许多植物也随之分布到中纬度许多地方。后来间冰期来到,冰川北退,气候变暖,它们就在气候寒冷的高山顶部保存下来,形成目前的分布。
有些间断分布很难从生物的繁殖能力、散布方式和生境特点等方面圆满解释,必须考虑历史因素。例如假山毛榉属(Nothofagus)目前的分布范围是南半球的智利南端、新西兰、澳大利亚、新喀里多尼亚和新几内亚,但非洲没有。有人研究了这个属的散布特征得出结论:它的种子不适于跳跃式散布,因而不能越过广阔的咸水海洋,它现在的不连续分布是冈瓦纳古陆破裂产生的结果。A.L.韦格纳的大陆漂移学说,随着新近板块理论的出现已得到普遍承认。按照大陆漂移说,大陆曾经是连接在一起的,后来逐渐裂开。在中生代的某个时候,全部世界的陆地组成两块大的大陆:劳亚古陆和冈瓦纳古陆。劳亚古陆由现在的北美和欧亚大陆组成;冈瓦纳古陆由现在的南极大陆、澳大利亚、南美、非洲和印度组成。这两块古陆中间隔着古地中海。后来劳亚古陆和冈瓦纳古陆破裂成较小的部分。冈瓦纳古陆的碎块向外漂移开,就形成现在南半球的几块陆地。假山毛榉是冈瓦纳属,它的分布限于冈瓦纳古陆,虽然现在在南极大陆没有假山毛榉的分布,但南极大陆上有两个地点发现了假山毛榉的花粉化石,所以该属今天的间断分布应是冈瓦纳古陆破裂的结果形成的。
地球的生物地理分区
如果把地球不同部分的植物和动物种类进行对比,就会发现彼此有明显的差别,两地相距越远差别越大。虽然可能有些分类单位彼此共有,但通常是许多分类单位只出现在某一地区,而不出现在另一地区。这就有可能把地球划分为不同的植物地理区域(见彩图)和动物地理区域(见彩图),或者是同时考虑植物和动物的生物地理区。无论植物、动物或生物地理区域都是客观的存在,都是科学研究的实体。
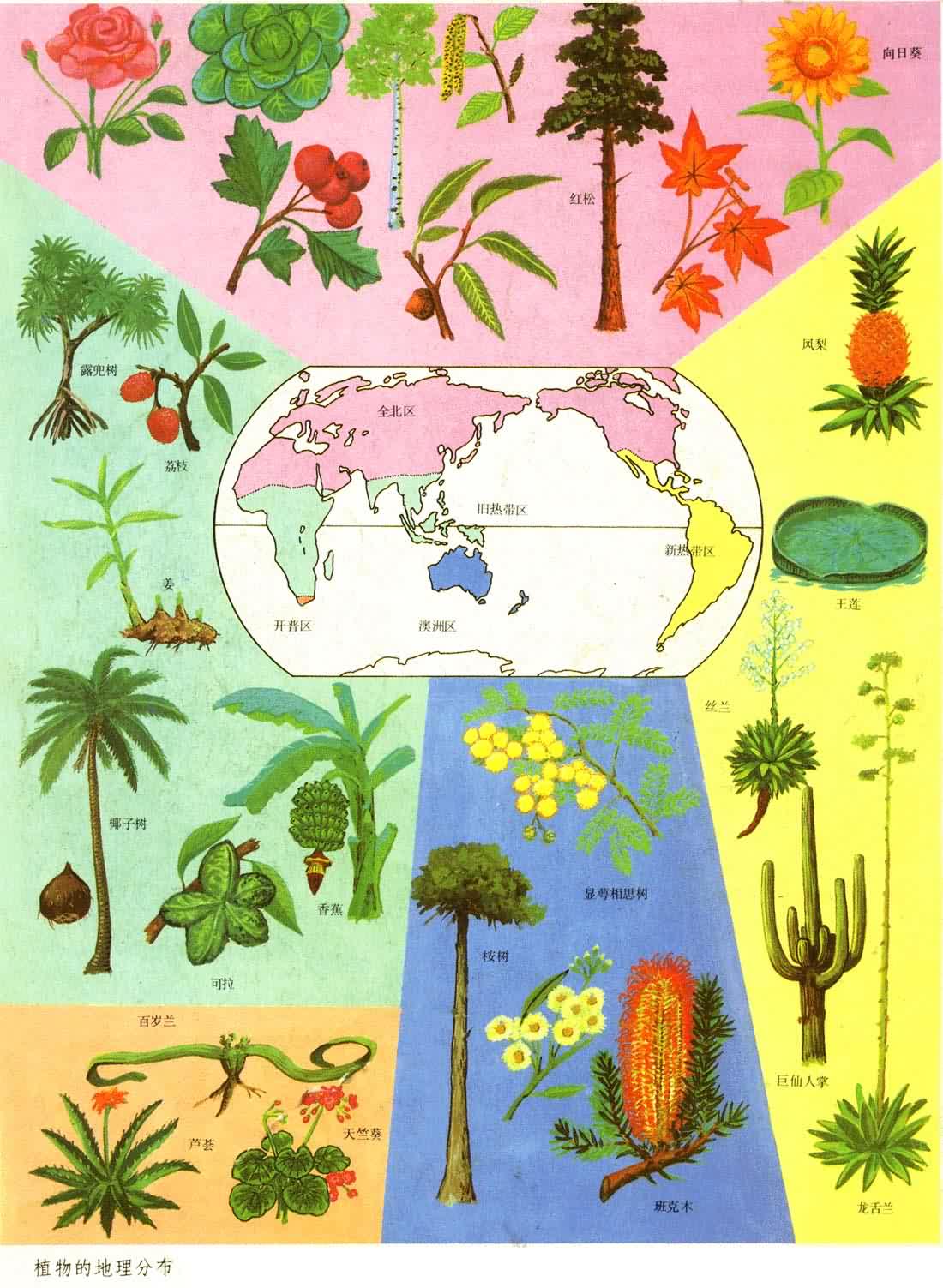
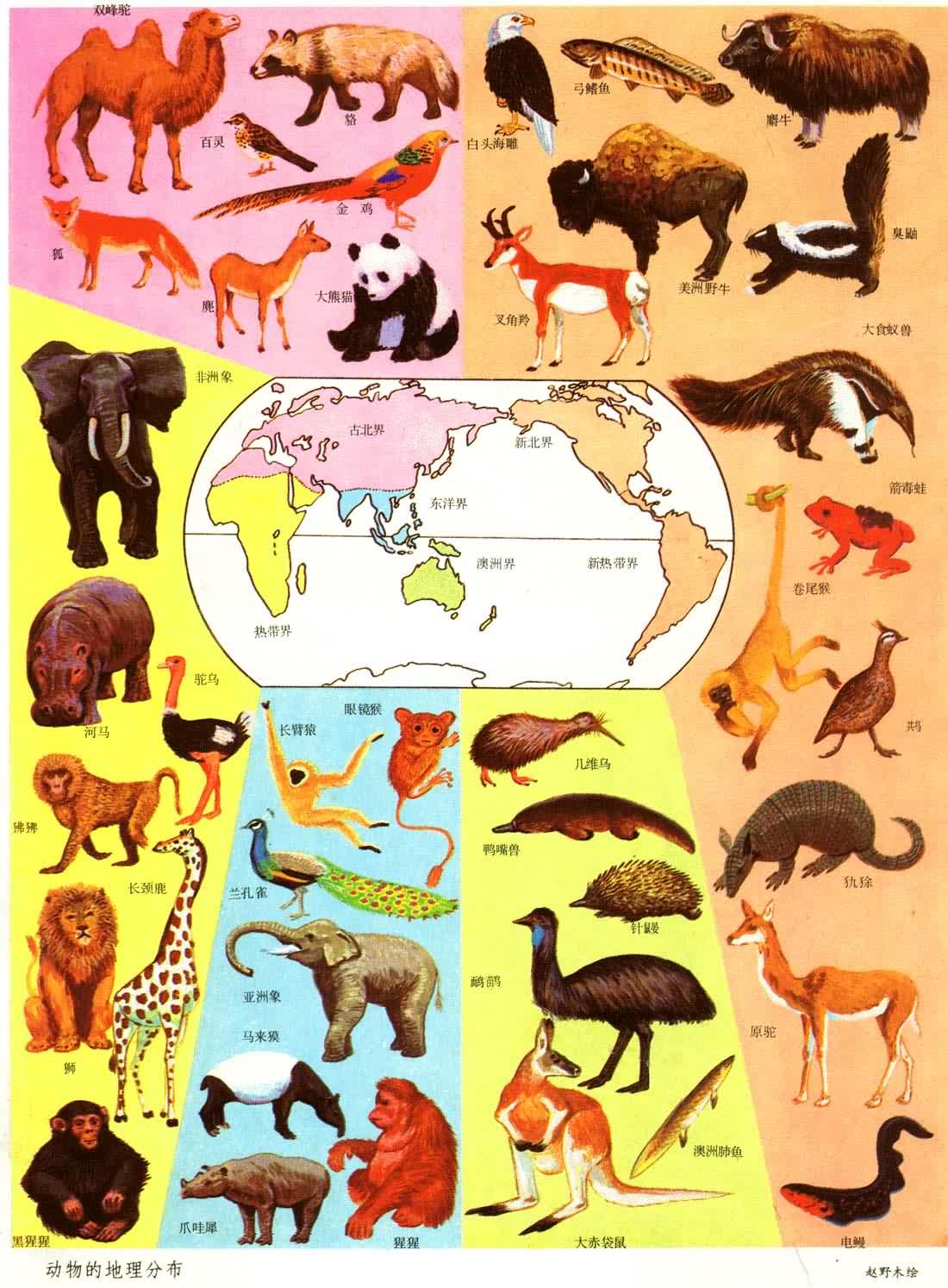
无论植物或动物,地理区域的划分必须首先根据常见的优势类群的分布。所以动物地理区域主要根据哺乳动物的分布来划分,而植物地理区域则根据被子(有花)植物的分布来划分。
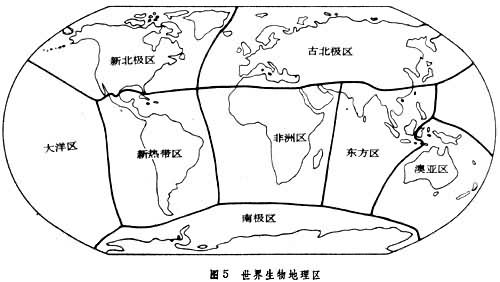
E.C.皮耶卢1979年对动物和植物给予同样的强调。把施密特的省结合成 8个“生物地理区”,即古北极区、新北极区、新热带区、非洲区、东方区、澳亚区、南极区,此外还包括一个大洋区(图5)。
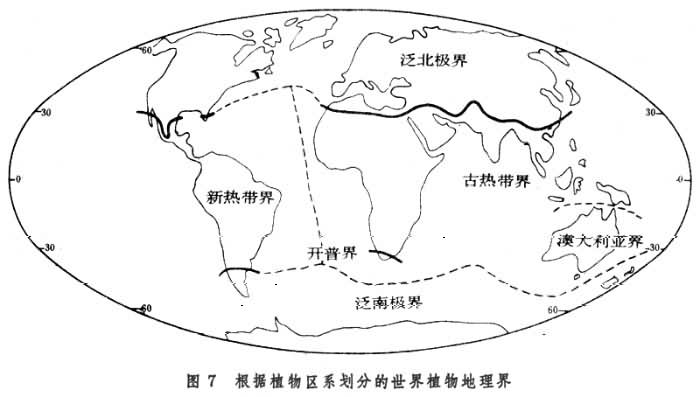
植物地理区域的划分从D.恩格勒,L.迪尔斯以来,通行分为3级:界、区和省。界的主要特征是具有特有科和亚科,并且属和种的特有率非常高;区的特征是有很高的属和种特有现象;省之内属的特有现象不显著,而以种的特有现象为特征。D.恩格勒将全世界分成 4个陆地植物区系界、1个海洋界;将陆地植物区系界分为29个区、102个省。近代较有代表的植物地理区域的划分,为英国植物学家R.古德和苏联植物学家А.Л.塔赫塔江所提出。古德将地球陆地植物区系分为6个界、37个区、120个省(图6);
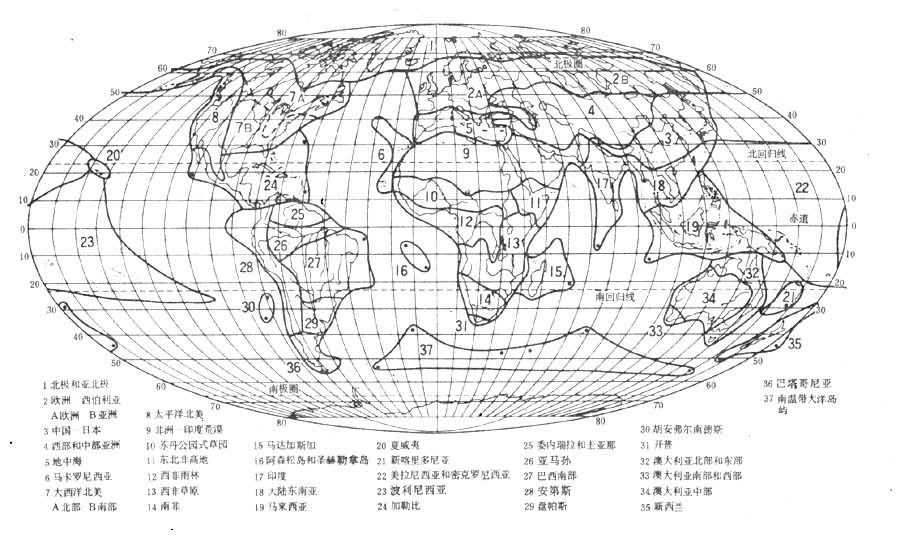
А.Л.塔赫塔江则分为6个界、34 个区、147个省。可见在植物地理学中采用的界的划分数目不大,划分的结果也比较一致,普遍同意将陆地植物区系划分为 6个界──泛北极界、古热带界、新热带界、开普界、澳大利亚界和泛南极界(图7)。
动物地理区域的划分,从A.R.华莱士以来公认的方案也是将地球陆地分为6个动物地理界,每一个或多或少包括一个主要大陆,并且互相间被海洋或山脉分开。它们是:古北界、新北界、新热界、非洲界、东洋界和澳洲界。
较新的一个动物地理划分方案是K.P.施密特1954年提出的,他区分出4个不同等级的单位:界、区、亚区和省。他一共划分出3个界(北界、新界、南界)、5个区(古热区、全北区、新热区、澳大利亚区、大洋区)、13个亚区和32个省。
海洋可以分为两个界:大陆架和深海。200米深等深线是大陆架和深海的分界线。这是光透入的下限。在此限度以上的海洋生物能进行光合作用。深海从来没有成为过陆地,那里动物区系分布简单,在世界范围内比较一致,具有广泛分布的世界性科。而大陆架实际是大陆的一部分,沉没在陆缘海之下,动植物区系较为复杂多样。