[拼音]:jianduan pinghenglun
[外文]:punctuated equilibrium
认为进化过程是突变与渐变交替出现的一种生物进化学说。1972年由美国古生物学家N.埃尔德雷奇和S.J.古尔德提出后,在欧美流传颇广。它与传统的进化论区别有三:
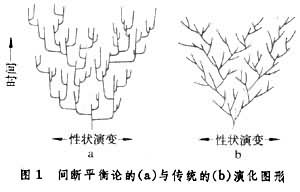
(1)传统学说强调进化是物种在自然选择下的渐进演变过程,在时间(纵向)-性状演变(横向)坐标上呈斜线的形式(图1b);间断平衡论则认为进化是突变与渐变的结合。强调大多数物种的形成是在地质上可忽略不计的短时间内完成的,这个迅速的过程叫种形成,在上述坐标上表现为接近水平的横线。物种形成后,在选择作用下发生的十分缓慢的变异,叫线系渐变,在上述坐标上表现为接近于垂直的纵线。整个进化呈现折线的图形(图1a)。
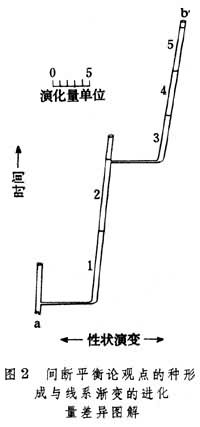
(2)传统进化论认为进化量(即生物种系在一段时间内的性状演变总量)是渐进变异逐渐积累的总和,线系渐变是进化的主流。间断平衡论则认为虽然渐变也可造成变异,并积累形成新种,但其在总变异量中所占份额很小,种形成才是进化的主流。例如,将图1a中的一段放大成图2,可以看到从a种演化到b种,其进化量(ab在性状演变坐标上的投影)主要由水平横线(种形成)构成;种系渐变(1,2,3,4,5各种)虽也有水平分值,但由于它们接近于垂直,所以数值都不大(图2)。
(3)间断平衡论强调变异的随机性和地理隔离对种形成的必要性。它认为形成新种的原料是个体突变,突变是无定向的。只要对适应无害(中性),就可能闯过自然选择这一关而有可能形成新种。它又强调大多数新种是从父种地理分区边缘上被隔离的孤立小种群中形成的,因为在这孤立种群中产生的突变,不致因基因交流而失去特性,其中多数虽被淘汰,少数仍能被选择保留下来而形成新种。
依据
基因突变学说和常见的地理隔绝导致迅速成种的现象成为间断平衡论的主要依据。只要是处在关键性个体发育途径上的基因,一个基因的一个突变即足以造成新种。在多细胞生物(包括人在内)中,每一代中每个基因产生突变的或然率是10-4~10-9,其中间值为十万分之一(10-5)。一个普通生物个体约有30000对基因,即每一代中平均每10个个体中有 6个基因突变。当然其中只有极少一部分能通过自然选择考验,在以后世代中稳定下来,形成新种。因此,有的遗传学家认为新种是可以在几个生殖世代中形成的。种群中的个体突变在化石中不易找到。曾经报道过美国阿拉巴马州始新世的小帘心蛤种群中,少数个体左右壳的牙系与正常个体恰巧相反的情况。
下面举一个由于隔离而迅速成种,甚至成属的例子。2~3万年前,美国西部的河湖中广泛分布着一种鳉鱼。现在这里只有互相隔绝的残余小湖及泉。在加州死谷区的几个孤立池、泉中分别发现了 4个不同种,显然是在这段时间中由原来种形成的。特别是产于某温泉中的一个种──魔鬼泉鳉鱼,其性状演变几乎已经达到可建立新属的程度。换言之,新属可以在2~3万年的短时间形成。有的例子说明,自然界新种的形成可短到250年。
争议
间断平衡论问世后,赞成方面的,有人引用小泽(1975)对多壁小鳞(虫筳)初房直径演变所作的统计。在该种延续的1500万年内,直径从平均200微米渐增至550微米,仍不超出种的范围。说明在不隔离情况下,单纯的种系演变是非常缓慢的,不易形成新种。在东非肯尼亚上新世至更新世,有一段300~500万年的地层,在190个化石点采集后,对常见的13个软体动物种3300块标本,每个作了平均18种度量,用计算机作数理统计,根据其形态演变情况得出三点结论:
(1)所有种系在长时期中形态保持稳定。例如对单色环棱螺共采集了49个层位,在绝大多数层位中其形态保持不变;
(2)在 3个层位上有急剧的变异,产生新种。这 3个是凝灰岩层位,各代表时间间隔 5000~50000年,所以成种作用是迅速的;
(3)无论旧种和新种,变异幅度都很小,只有在从旧种到新种的过渡种群中才产生大幅度的不定变异。这个资料显然支持间断论,即种形成过程是迅速的突变;种一旦形成,性状长期稳定;只有在种形成(属或更高级别的形成)过程中,才产生大量不定变异(W.C.威廉森,1981)。
反对间断平衡论的例子如G.金格里奇1974年统计北美兔猴各个种下第一臼齿长宽积的变化,指出种与种之间臼齿面积是渐变的,甚至新属形成也是渐变的。M.马尔姆格伦等(1982)对新西兰附近深海钻探发现的中新世至更新世某有孔虫属的各种形态变化作了统计,发现这些变化大都是渐变的。
目前古生物学界多数承认种形成过程可以是快速突发的也可以是渐变的。如K.考夫曼1978年对近 100个白垩纪双壳类种系的演化型式研究表明,有渐变式的,网格式的(种的各种群在演变的环境中通过反复的隔离,重新融合、杂交,最后演化的新种)和间断突变式的。这取决于环境压力变化的强度和速度以及种的适应宽度。
意义、展望
间断平衡论以一种新的哲学观点来看待生物进化史,向古生物学和生物学的传统观点提出了挑战。
(1)间断平衡论较合理地解释了化石记录。按照传统的渐进进化观点,化石记录应当是一个循序渐进的连续过程。但是百余年积累下来的记录却不是这样,在连续地层(也就是连续的时间序列)中,新门类往往是突然出现的,找不到其祖先的蛛丝马迹。现在按照间断平衡论,既然新种、新属可以突变形成,那么由它们组成的新门类(科、目,以至纲、门)也可以在地质上可忽略不计的时间内形成。生物界的突变,跃进到新阶段,可以在没有地质间断的层序中发生。在中国南方,古生界和中生界之间往往是连续沉积的,看不到长时间的间断。但是古生代和中生代的生物面貌却是突变的,无论菊石、腕足类、双壳类、有孔虫类、牙形石、三叶虫类、珊瑚类等都有突然的变化或甚至绝灭。为了解释这种貌似矛盾的现象,曾经引进了“拟整合”的概念,即自然界存在着一种从岩性结构上看不出,只能根据化石记录缺失来判断的地质间断。现在根据间断平衡论,生物界突变可以在地质上忽略不计的时间中完成,不再必需一个长期间断来解释生物界的突变了。在前寒武纪-寒武纪界面上同样存在层序整合而具壳生物突然大量出现的现象。过去一种解释是,这些门类在前寒武纪早已存在,只是因为尚未形成硬壳,所以缺少化石记录,这解释有其矛盾之处。C.克劳德1948年指出:腕足动物如果不具硬壳,就不叫腕足动物,而属于帚虫动物。许多带壳动物不大可能有很长的前寒武纪历史。根据突变论,可以假设前武纪-寒武纪之交,以迅速的种形成向各个适应方向产生新类型,然后固定成新门类,并不需要每一门类有很长的缺失化石记录的历史。再推而广之,过去人们碰到处于两个演化阶段的古生物,总是努力设法去发现其中间类型以证明演化的渐进性。如果中间环节找不到,则归咎于化石记录的不完整。尽管如此,绝大多数演化阶段都是找不到中间型的。现在看来,这可能不是化石记录的缺失,而是正常的现象。通常是由一个祖先类型向各个方向产生突变。这些突变大部分是不成功的,少数在成功的方向上经过一系列的突变──种形成,就可以选成一新的类型。由于突变是很迅速的,在地层中不易记录;更由于大种群中的突变常因基因交流而失去,往往只有在小范围分布的小种群中突变才能成功,所以地理分布亦很狭。因此中间环节是很难找到的。
(2)间断平衡论展示了一幅不同于传统达尔文主义所描绘的生物演化图,以菊石的演化为例,这个门类在晚泥盆世至早石炭世、早三叠世、早侏罗世有几个爆发期,在很短的时间内向各个生态领域演化,形成许多新科(这种现象叫辐射演化)。这个新门类形成以后在石炭二叠纪,三叠纪,侏罗白垩纪的较长时间内保持相对平衡。突变(新门类的爆发形成)与渐变(已形成门类的相对稳定发展)交替出现,而每一次突变都是继前一阶段门类的绝灭之后,由其中一个小分支爆发造成的。这样,爆发期造成的阶段性与其后渐变演化造成的连续性,就构成菊石演化的基本图案。在别的门类中也同样呈现这种阶段性与连续性结合或间断-平衡的现象。
(3)目前进化论方面的间断平衡论,分类学方面的分支系统学,生物地理学方面的替代分化模式三者结合在一起,强烈地冲击着达尔文主义问世以来形成的现代古生物学概念。从哲学上来说,它们强调的突变与渐变相结合而以突变为主,以严格的系统发育关系作为分类学的基础等──有正确的方面,对于近百年来流行的渐变观点是一个否定。这一否定与板块学说及其他学科的新学说一起,正在对地球科学及生物科学产生巨大影响。
间断平衡论的一些极端支持者倾向于排斥其他可能的进化途径,并且在支持者内部使用一套特有的术语和概念。在社会上,神学界利用间断平衡论与特创论形式上的某些相似,宣传上帝的创造,否认进化。因此,新理论要存在和发展下去,还须经历去芜存精的长期洗炼过程。