[拼音]:chongzu DNA jishu
[外文]:recombinant DNA technique
又称遗传工程,在体外重新组合脱氧核糖核酸(DNA)分子,并使它们在适当的细胞中增殖的遗传操作。这种操作可把特定的基因组合到载体上,并使之在受体细胞中增殖和表达。因此它不受亲缘关系限制,为遗传育种和分子遗传学研究开辟了崭新的途径。
广义的遗传工程包括细胞水平上的遗传操作(细胞工程)和分子水平上的遗传操作,即重组DNA技术(有人称之为基因工程)。狭义的遗传工程则专指后者。
简史
重组DNA技术来源于两个方面的基础理论研究──限制性核酸内切酶(简称限制酶)和基因载体(简称载体)。限制酶的研究可以追溯到1952年美国分子遗传学家S.E.卢里亚在大肠杆菌中所发现的一种所谓限制现象──从菌株甲的细菌所释放的噬菌体能有效地感染同一菌株的细菌,可是不能有效地感染菌株乙;少数被感染的菌株乙的细菌所释放的同一噬菌体能有效地感染菌株乙可是不能有效地感染菌株甲。经过长期的研究,美国学者W.阿尔伯在1974年终于对这一现象提出了解释,认为通过噬菌体感染而进入细菌细胞的 DNA分子能被细菌识别而分解,细菌本身的DNA则由于已被自己所修饰(甲基化)而免于被分解。但有少数噬菌体在没有被分解以前已被修饰了,这些噬菌体经释放后便能有效地感染同一菌株的细菌。被甲(或乙)这一菌株所修饰的噬菌体只能有效地感染甲(或乙),而不能有效地感染乙(或甲),说明各个菌株对于外来DNA的限制作用常常是专一性的。通过进一步的研究发现这种限制现象是由于细菌细胞中具有专一性的限制性核酸内切酶的缘故。
重组DNA技术中所用的载体主要是质粒和温和噬菌体(见转导)两类,而在实际应用中的载体几乎都是经过改造的质粒或温和噬菌体。英国微生物遗传学家W.海斯和美国微生物遗传学家J.莱德伯格等在1952年首先认识到大肠杆菌的 F因子(见细菌接合)是染色体外的遗传因子。1953年法国学者P.弗雷德里克等发现大肠杆菌产生大肠杆菌素这一性状为一种染色体外的大肠杆菌素因子所控制。1957年日本学者发现了抗药性质粒。后两类质粒都是在遗传工程中广泛应用的质粒。
重组DNA技术中广泛应用的噬菌体是大肠杆菌的温和噬菌体λ,它是在1951年由美国学者E.莱德伯格等发现的。
到 70年代初,生物化学研究的进展也为重组DNA技术奠定了基础。1972年美国的分子生物学家P.伯格等将动物病毒 SV40的DNA与噬菌体P22的DNA连接在一起,构成了第一批重组体 DNA分子。1973年美国的分子生物学家 S.N.科恩等又将几种不同的外源DNA插入质粒pSC101的DNA中,并进一步将它们引入大肠杆菌中,从而开创了遗传工程的研究。
步骤和技术路线
重组DNA技术一般包括四步:
(1)产生DNA片段;
(2)DNA片段与载体DNA分子相连接;
(3)将重组DNA分子导入宿主细胞;
(4)选出含有所需要的重组体 DNA分子的宿主细胞。在具体工作中选择哪条技术路线,主要取决于基因的来源、基因本身的性质和该项遗传工程的目的(见图)。
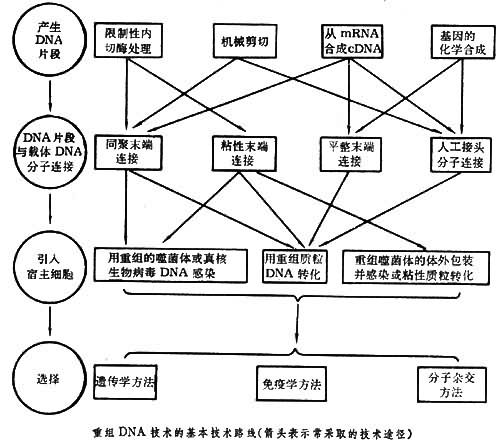
重组DNA片段的取得
主要的方法有:
(1)利用限制酶取得具有粘性末端或平整末端的 DNA片段;
(2)用机械方法剪切取得具有平整末端的DNA片段,例如用超声波断裂双链DNA分子;
(3)经反向转录酶的作用从mRNA获得与mRNA顺序互补的DNA单链,然后再复制形成双链DNA(cDNA)。例如人的胰岛素和血红蛋白的结构基因都用这方法获得。这样获得的基因具有编码蛋白质的全部核苷酸顺序,但往往与原来位置在染色体上的基因在结构上有区别,它们不含有称为内含子的不编码蛋白质的间隔顺序(见基因);
(4)用化学方法合成 DNA片段。从蛋白质肽链的氨基酸顺序可以知道它的遗传密码。依照这密码用化学方法可以人工合成基因。
DNA片段和载体的连接
DNA片段和载体相连接的方法主要有四种:
(1)粘性末端连接,每一种限制性核酸内切酶作用于 DNA分子上的特定的识别顺序,许多酶作用的结果产生具有粘性末端的两个 DNA片段。例如来自大肠杆菌(Escherichia coli)的限制酶 EcoRI作用于识别顺序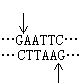 (↑指示切点),产生具有粘性末端
(↑指示切点),产生具有粘性末端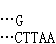 和
和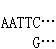 的片段。把所要克隆的DNA和载体DNA用同一种限制酶处理后再经 DNA连接酶处理,就可以把它们连接起来。
的片段。把所要克隆的DNA和载体DNA用同一种限制酶处理后再经 DNA连接酶处理,就可以把它们连接起来。
(2)平整末端连接,某些限制性内切酶作用的结果产生不含粘性末端的平整末端。例如来自副流感嗜血杆菌(Hemophilus parainfluenzae) 的限制酶Hpal作用于识别顺序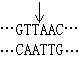 而产生末端为
而产生末端为  的DNA片段。用机械剪切方法取得的DNA片段的末端也是平整的。在某些连接酶(例如感染噬菌体T4后的大肠杆菌所产生的DNA连接酶) 的作用下同样可以把两个这样的DNA片段连接起来。
的DNA片段。用机械剪切方法取得的DNA片段的末端也是平整的。在某些连接酶(例如感染噬菌体T4后的大肠杆菌所产生的DNA连接酶) 的作用下同样可以把两个这样的DNA片段连接起来。
(3)同聚末端连接,在脱氧核苷酸转移酶(也称末端转移酶)的作用下可以在DNA的3′羧基端合成低聚多核苷酸。如果把所需要的 DNA片段接上低聚腺嘌呤核苷酸,而把载体分子接上低聚胸腺嘧啶核苷酸,那么由于两者之间能形成互补氢键,同样可以通过DNA连接酶的作用而完成DNA片段和载体间的连接。 ④人工接头分子连接,在两个平整末端 DNA片段的一端接上用人工合成的寡聚核苷酸接头片段,这里面包含有某一限制酶的识别位点。经这一限制酶处理便可以得到具有粘性末端的两个DNA片段,进一步便可以用DNA连接酶把这样两个DNA分子连接起来。
导入宿主细胞
将连接有所需要的 DNA的载体导入宿主细胞的常用方法有四种:
(1)转化,用质粒作载体所常用的方法。
(2)转染(见转化),用噬菌体DNA作载体所用的方法,这里所用的噬菌体 DNA并没有包上它的外壳。
(3)转导,用噬菌体作载体所用的方法,这里所用的噬菌体DNA被包上了它的外壳,不过这外壳并不是在噬菌体感染过程中包上,而是在离体情况下包上的,所以称为离体包装。
(4)注射,如果宿主是比较大的动植物细胞则可以用注射方法把重组DNA分子导入。
选择
用以上任何一种方法连接起来的 DNA中既可能包括所需要的 DNA片段,也可能包括并不需要的片段,甚至包括互相连接起来的载体分子的聚合体。所以接受这些 DNA的宿主细胞中间只有一小部分是真正含有所需要的基因的。一般通过 3种方法可以取得所需要的宿主细胞:
(1)遗传学方法,对于带有抗药性基因的质粒来讲,从被转化细菌是否由敏感状态变为抗药的状态就可以知道它有没有获得这一抗药性质粒。一个抗药性基因中间如果接上了一段外来的 DNA片段,就使获得这一质粒的细菌不再表现抗性。把一个带有两个抗性基因氨苄青霉素抗性和四环素抗性的质粒 pBR322用限制酶 Bam HI处理,由于Bam HI的唯一的识别位点是在四环素抗性基因中,所以经同一种酶处理的DNA分子片段就可以连接在这一基因中间。在被转化的细菌中选择只对氨苄青霉素具有抗性而对四环素不具抗性的细菌,便可以获得带有外来 DNA片段的载体的细菌。这是一种常用的遗传学方法。
(2)免疫学方法和分子杂交方法,当一个宿主细胞获得了携带在载体上的基因后,细胞中往往就出现这一基因所编码的蛋白质,用免疫学方法可以检出这种细胞。分子杂交的原理和方法同样可以用来检测这一基因的存在(见分子杂交、基因文库)。
基因表达
在构建重组体 DNA分子和选择宿主细胞时,还须考虑外源基因表达的问题。就是说要求外来的基因在宿主细胞中能准确地转录和翻译,所产生的蛋白质在宿主细胞中不被分解,而且最好还能分泌到细胞外。为了使外源基因表达,需要在基因编码顺序的5'端有能被宿主细胞识别的启动基因顺序以及核糖体的结合顺序。两种常用的方法能用来使外源基因在宿主细胞中顺利地表达:
(1)在形成重组体DNA分子时在载体的启动基因顺序和核糖体结合顺序后面的适当位置上连接外源基因。例如将兔的β-珠蛋白基因或人的成纤维细胞干扰素基因分别连接到已经处在载体上的大肠杆菌乳糖操纵子的启动基因后面,便能使它们在大肠杆菌中顺利地表达;
(2)将外源基因插入到载体的结构基因中的适当位置上,转录和翻译的结果将产生一个融合蛋白。这种融合蛋白质被提纯后,还要准确地将两部分分开,才能获得所需要的蛋白质。在早期的遗传工程研究中,生长激素释放抑制因子和鼠胰岛素基因的表达都是通过将它们连接在β-半乳糖苷酶基因中的方式实现的。
应用
发酵工业
用大肠杆菌生产人的生长激素释放抑制因子是第一个成功的实例。在 9升细菌培养液中这种激素的产量等于从大约50万头羊的脑中提取得到的量。这是把人工合成的基因连接到小型多拷贝质粒pBR322上,并利用乳糖操纵子β-半乳糖苷酶基因的高效率启动子,构成新的杂种质粒而实现的。现在胰岛素、人的生长激素、人的胸腺激素 α-1、人的干扰素、牛的生长激素、乙型肝炎病毒抗原和口蹄疫病毒抗原等都可用大肠杆菌发酵生产,其中有的还可在酵母或枯草杆菌中表达,这就为大规模的工业发酵开辟了新的途径。还有些很重要的基因,如纤维素酶的基因等也已在大肠杆菌中克隆和表达。
利用遗传工程手段还可以提高微生物本身所产生的酶的产量。例如可以把大肠杆菌连接酶的产量提高 500倍。
理论研究
应用重组DNA技术可以克隆和扩增某些原核生物和真核生物的基因,从而可以进一步研究它们的结构和功能。重组DNA技术的成就和提出的问题促进了遗传学、生物化学、微生物学、生物物理学和细胞学等学科的发展,并且有助于这些不同学科的结合。目前正在形成一门新兴的学科──生物工艺学或生物工程学,就是这种趋势的反映。
动植物育种和基因治疗
已经有一些研究工作明确地预示着重组DNA技术在这些方面的潜力。例如把来自兔的β-血红蛋白基因注射到小鼠受精卵的核内,再将这种受精卵放回到小鼠输卵管内使它发育,在生下来的小鼠的肝细胞中发现有兔的β-血红蛋白基因和兔的β-血红蛋白。还有人把包括小鼠的金属巯基组氨酸三甲基内盐 Ⅰ(metallothioneine Ⅰ)基因的启动子及大鼠生长激素结构基因的 DNA片段注射进小鼠受精卵的前核中,由此发育得来的一部分小鼠由于带有可表达的大鼠生长激素基因,所以明显地比对照鼠长得大。这些实验结果为基因治疗展现了可喜的前景。固氮的功能涉及17个基因,分属 7个操纵子,现在已能把它们全部引入酵母菌,而且能正常地复制,不过还没有能使这些基因表达。改造玉米胚乳蛋白质而使人畜营养必需的赖氨酸和色氨酸成分增加的工作也在着手进行。大豆的基因已能通过Ti质粒引入向日葵。因此,可以预期随着时间的推移在能源、农业、食品生产、工业化学和药品制造等方面都将会取得巨大的成果。
- 参考书目
- R.W.Old,S.B.Primrose,Principles of Gene Manipulation,University of California Press,1980.