[拼音]:fanshe
[外文]:reflex
动物通过中枢神经系统对刺激的一种应答式反应。这种反应的发生有赖于反射弧的完整,是一种刻板的简单动作,主要是肌肉收缩或腺体分泌。反射在具中枢神经系统的动物中普遍存在。机体通过反射来控制和调节体内各种生理过程,使它们相互协调,也使机体对环境的各种变化发生适应性反应,保证了机体与外环境的统一。
反射的概念
首先由法国哲学家R.笛卡儿提出。他注意到机体对于一些环境刺激具有规律性反应。例如,异物碰到角膜即引起眨眼。他借用了物理学中反射的概念,认为动物的活动像光线投射到镜子上被反射出来一样,即机体受到的刺激和发生的反应有必然的因果关系。用实验分析的方法研究脑和脊髓并阐明反射规律的,是英国的C.S.谢灵顿以及俄国的И.M.谢切诺夫、И.П.巴甫洛夫。谢灵顿对中枢神经系统低级部位(包括脑干、脊髓)的反射作了详细研究,阐明了反射的基本规律。但是他只把外界刺激引起机体的非随意动作叫作反射,并认为皮层以下的神经核团是各级反射的中枢。一般不把反射概念应用到神经系统高级部位(大脑)的活动中。谢切诺夫将反射概念应用于人脑的活动。以后巴甫洛夫在谢切诺夫思想影响下,进一步研究大脑皮层的功能。他通过利用狗所做的著名试验把唾液腺的所谓心理性分泌理解为一种特殊的反射──条件反射,并客观地研究了这种活动的发生与消退等规律,提出了条件反射学说,即高级神经活动学说。此项研究倾向于对反射的神经回路及递质机制作更深入的分析。
种系发生与个体发生
单细胞动物无神经系统,对刺激的反应都由原生质来完成。腔肠动物开始有神经细胞,并形成网状,但无中枢和外周之分,当受到刺激时则引起全身反应。环节动物和节肢动物具有链状神经系统,咽部背侧一对神经节特别大,有人也把它叫做脑,这些动物有完整的反射弧(图1),对内、外环境的刺激开始有了简单的反射活动。
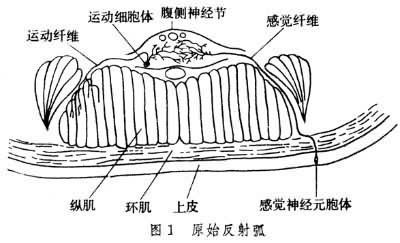
脊椎动物的中枢神经系统的前端为真正脑,后端为脊髓;外周神经由脑神经和脊神经组成,其中哺乳动物出现了高度发展的大脑皮层。在高等动物以至人类,大脑皮层已成为控制整个机体活动的高级部位。中枢神经系统各结构在功能上有高级与低级之分,低级中枢可完成一些基本的、简单的反射,在高级中枢调节下反射则更加复杂而完善。
人胚发育至第3周即开始形成神经板,以后发展为神经管,再由神经管头端膨大发展为脑的不同部分,而由神经管的尾端发展成脊髓。由于胎儿神经系统的基本结构已经形成,因此初生儿就可完成一些最简单的非条件反射,如吸吮反射、吞咽反射、排尿反射、排便反射和屈肌反射等。以后由于机体不断接触外界环境,经过训练可逐渐形成越来越多的条件反射。在个体成长过程中,不断建立各种条件反射,使反射的数量越来越多,反射的形式也越来越复杂。
反射弧
反射活动的结构基础。是机体从接受刺激到发生反应的过程中兴奋在神经系统内循行的整个路径。反射一般都需要完整的反射弧来实现。一个完整的反射弧由感受器、传入神经、神经中枢、传出神经、效应器5个基本部分组成(图2)。
 感受器
感受器
能将内外环境的刺激转变为神经冲动的特殊结构;是反射活动的起始处。
传入神经感觉神经元的突起。将感受器的神经冲动传导到神经中枢。
神经中枢中枢神经系统内参与某一反射活动的神经元群,是反射弧的中枢整合部分。例如参与瞳孔对光反射的中枢结构叫做对光反射中枢。
简单反射的中枢范围比较局限,在神经中枢内传入神经元可直接与传出神经元形成单突触联系;多数反射在传入与传出神经元之间要经过一个以上中间神经元的接替,形成多突触反射。
传出神经运动神经元的轴突。将反射中枢发生的神经冲动传到效应器。
在反射活动中,运动神经元既接受传入神经或中间神经元传入冲动的影响,又接受各级中枢下行纤维的控制。因此,它在反射传出的通路上成为最后公用的、到效应器传出路径,谢灵顿叫它做最后公路。
效应器发生应答反应的器官或组织,包括骨骼肌、心肌和平滑肌,各种外分泌腺和一些内分泌腺等。
反射弧的5个组成部分中任何部分的中断,都会使反射消失。在某些反射活动中,传出神经首先作用于某些内分泌腺,使该腺体释放激素经血液转运,最后作用于效应器。这种有内分泌腺参与的反射活动,其效应的出现往往比较缓慢,但影响比较广泛而持久。
反射的分类
根据反射的特点,有不同的分类法。
(1)按反射形成的特点可分为非条件反射和条件反射两大类:前者是动物生来就有的,无需后天训练的反射,它是动物在种系进化过程中建立和巩固起来的,可再遗传给后代。非条件反射的反射弧是固定的,其数目有限,如牵张反射、瞳孔对光反射等。后者是动物在后天的个体生活中经过学习和训练而获得的,是反射的高级形式。如果动物的生活条件发生改变则已形成的条件反射会消退,并可重新形成新的条件反射。因此,条件反射的反射弧不是固定不变的,其形式是多样的、数目是无限量的。它使动物对于千变万化的外界环境具有更大的适应性。
(2)按感受器的部位不同可分为由外感受器引发的外感受性反射和由内脏或本体感受器引起的内感受性反射。
(3)按效应器的性质可分为躯体反射和内脏反射。前者的效应是骨骼肌的活动,如姿势反射等;后者的效应包括心血管、胃肠道、支气管平滑肌、膀胱与输尿管运动的变化以及腺体的分泌等。
(4)按反射的生物学意义可分为防御反射或保护性反射,摄食与消化食物的食物反射以及与延绵种族有关的性反射。
(5)按反射弧中突触的数目多少可分为单突触反射与多突触反射,前者是通过单突触反射弧的反射。如膝跳反射,即膝关节下方的肌腱受到扣击后,肌肉内的感受器(肌梭)受到刺激产生的神经冲动,沿传入神经进入脊髓直接通过与运动神经元形成的突触引起后者兴奋,产生一次肌肉收缩的过程。后者是通过两个以上突触的反射弧发生的反射。体内多数反射均为多突触反射。如屈肌反射,即指疼痛或伤害性刺激作用于皮肤或皮下组织时,可引起受刺激肢体的屈肌收缩,伸肌宽息,从而使该肢体缩回的反射,就是多突触反射。
(6)从临床的角度考虑,可将反射分为生理反射和病理反射。前者是正常生命过程中出现的各种反射,后者则是在神经系统的某些部位发生病变后出现的异常反射。瞳孔对光反射即属生理反射,而锥体系破坏后出现的踝阵挛和巴彬斯基征等属病理反射。某些反射常是医生判断某一部位神经系统的功能是否正常的指标。
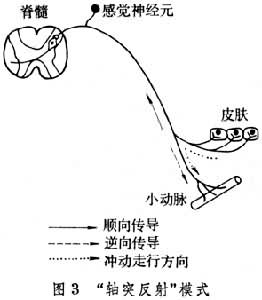
还有一些特殊种类的“反射”,如“局部反射”与“轴突反射”等。严格地说,它们都不是真正的反射,即指不需要中枢神经系统的参与就可完成的反射。如低等动物经过神经网与神经节的某些反应而形成的即属“局部反射”。高等动物的肠壁内的神经节和神经丛也可完成一定的“局部反射”(见肠神经系统)。“轴突反射”是当皮肤感受器受到刺激时,可引起受刺激部位周围的皮肤发生红晕的现象。是皮肤感受器兴奋时,神经冲动沿传入神经元突起的其他分支逆向传到外周的皮下血管,并未经过中枢而直接引起皮肤血管舒张的反应(图3)。
反射活动的基本特征
在反射活动中神经冲动必须经过反射弧的中枢部分。中枢神经元之间并无原生质相连,它们之间的信息传递主要靠神经元相接触的突触或缝隙连接来完成。因此,信息通过中枢内的传递已经不同于外周的传导,再加上神经元之间连接的方式复杂多样,因此,使反射活动具有与外周传导不同的许多特征:
单向传布神经冲动在中枢部分传布时,只能由传入神经元向传出神经元的方向进行,不能逆传。这种单向传布的特征由中枢内突触传递的特性决定。因此,在反射弧中兴奋只能从脊髓背根传入,由腹根传出,而不能相反,即背根是感觉性的,腹根是运动性的,这个规律叫做贝尔-马让迪定律。
中枢的兴奋过程与抑制过程是中枢神经活动的两种基本过程。两者都发生在突触处。当一神经元兴奋时,通过突触传递可能对后续的神经元发生两种影响:兴奋或抑制(见突触)。
除最简单的反射弧由两神经元组成外,复杂的反射弧则由许多神经元组成,每个神经元还可同时接受许多兴奋性突触和抑制性突触的作用。而且这两种突触后电位的强度对比,又时刻受各种传入冲动的影响而发生变化。所以,反射活动是中枢兴奋和抑制过程互相作用的结果。如果中枢的兴奋占优势,则出现某一具体的反射;如果中枢的抑制占优势,则此反射减弱或不出现,叫做反射的抑制。
局限化与扩散在反射活动中,如果给予感受器的刺激强度适宜,一般只引起较局限的反射,而不引起广泛活动,叫做反射的局限化。如刺激过强,会引起广泛的活动,叫做反射的扩散。神经元之间的辐散式联系是反射扩散的结构基础。扩散的广度决定于刺激的强度与中枢不同的功能状态。例如,刺激动物一侧下肢趾端皮肤,只引起踝关节屈曲:如增加刺激强度,兴奋将在中枢内扩散,会使膝关节乃至髋关节也发生屈曲。进一步加强刺激,兴奋还可扩散到对侧中枢,引起对侧下肢伸直。因此,当一侧下肢受到损伤性刺激时,同侧屈肌中枢发生兴奋过程,引起该肢体屈曲,缩回,以避开刺激源;同时,兴奋扩散到对侧伸肌中枢,引起对侧伸肌反射,以支撑身体,维持一定的姿势。这样,受刺激一侧下肢屈曲,对侧下肢伸直,就会完成一个协调的姿势反射。
易化与阻塞在反射活动中,由单根纤维传入的一次冲动,不能使突触后神经元产生神经冲动,即不能引起反射效应。这是因为单根纤维传入的一次冲动所引起的兴奋性突触后电位很小,只能使神经元的兴奋性提高。但如有许多传入纤维同时传冲动到同一神经元,则在每个突触后膜上所产生的突触后电位可以叠加起来,这个现象叫做空间总和。它会使反射易化。
另一方面,冲动在反射弧的中枢部分传布时还可产生阻塞现象。例如,单独兴奋纤维A可引起9个突触后神经元兴奋,单独兴奋纤维B也可引起其他9个突触后神经元兴奋。由于纤维A和纤维B共同支配其中6个神经元,因此,当纤维A与纤维B兴奋时,只会有12个神经元而不是18个神经元发生兴奋。这个现象叫做阻塞。由此可知,在反射活动中,可由于中枢的易化而使反射活动加强,也可由于中枢的阻塞而使反射活动减弱。
中枢性延迟(中枢延搁)在反射活动中,从刺激开始到效应器产生反应之间的时间叫做反射时。它包括刺激引起感受器兴奋,产生神经冲动;神经冲动在传入神经和传出神经上传导以及神经冲动引起效应器发生反应所消耗的时间外,还包括冲动在中枢的突触处传递所消耗的时间。后者比冲动在相应长度的神经纤维上传导所需的时间要长得多。因此,叫做中枢性延迟或突触性延迟。据测定,兴奋通过一个突触所需要的时间为0.3~0.5毫秒,由此可知,在反射中枢内通过的突触数越多,中枢性延迟的时间越长。
后作用反射活动都由刺激引起,但当刺激停止以后,中枢兴奋并不立即消失,传出神经仍断续发放神经冲动,使反射活动延续一段时间,这一现象叫做反射的后作用或中枢兴奋的后放电。在一定范围内,刺激越强,或刺激作用时间越久,则后作用的延续时间也越长。中枢内神经元之间环路式的兴奋性突触联系是产生后作用的形态学基础。神经冲动经过网状联系中含有不同突触数目的许多侧枝,先后到达同一个传出神经元上,使传出神经元连续不断受到刺激而持续放电,是产生后作用的主要机制。
易疲劳性当重复快速刺激传入神经时,传出神经元最初的放电频率非常高,但经若干毫秒或数秒钟之后,放电频率便逐渐减少,这种就叫反射疲劳。反射疲劳发生在中枢的突触部位。因此,反射疲劳实质上就是突触传递的疲劳。突触疲劳的机制与突触前末梢递质的耗竭有关。据估计,如果神经递质的来源得不到及时补充,则突触前末梢所储备的神经递质仅够进行 1万次突触传递,其总量可在几秒钟内耗竭。
- 参考书目
参考文章
- 某些种类的海豚利用回声定位来发现猎物:它们发射出滴答的声音,然后接收水域中远处物体反射的回音。海洋生物学家推测这些滴答知识题库
- 当海面上空气各层的密度有较大差异时,远处的光线通过密度不同的空气层发生折射或全反射,海面上会出现远处物体的影像,这一现象知识题库
- 最近,英国科学家提出了一个大胆的设想,用500条由无线电控制的小船像空气中喷洒海水,以增强云层的反射能力,使热量远离地球知识题库
- 外抑制,指正在进行的条件反射由于新异刺激的出现而中止。按照上述定义,下列属于外抑制的是知识题库
- 玻璃幕墙的光反射并不能造成光污染知识题库
- 海市蜃楼是一种因光的折射而形成的自然现象.它是地球上物体反射的光经大气折射而形成的虚像知识题库
- 哪种降噪措施可以减弱反射声的强度?生产安全
- 为避免反射眩光,计算机屏幕应怎样摆放?生产安全
- 反射声比直达声最少延时多长时间就可听出回声?()生产安全
- 剖面测量的地震反射法SeismicReflectionProfiling地球科学