[拼音]:linzhi
[外文]:phospholipids
含磷酸的复合脂质。包括磷酸甘油酯(又称甘油磷酸酯)和鞘磷脂两类。生物体的重要组分,如动物的脑、肝、红细胞和卵黄等以及植物的种子含量较多,磷脂是细胞膜和各种细胞器(线粒体、内质网、细胞核、高尔基器、叶绿体等)膜的重要组分,几乎细胞所含有的全部磷脂都集中在生物膜中。生物膜的许多特性,如作为膜内外物质的通透性屏障,膜内外物质的交换,信息传递,神经脉冲的传导等都与磷脂和其他膜脂有关。磷酸甘油酯的主链是甘油,甘油的第三个羟基被磷酸酯化,另外两个羟基被脂肪酸酯化,磷酸基团又与各种结构不同的小分子化合物相连接。两个长碳氢链(脂肪酸链)具有非极性特性,甘油分子的第三个羟基与磷酸形成的酯键是有极性的;所以这类化合物是亲水脂两性分子。常见的磷酸甘油酯有磷脂酰胆碱(卵磷脂)、磷脂酰乙醇胺(脑磷脂)等。鞘磷脂的主链是鞘氨醇(含氨基的长链醇类化合物),脂肪酸以酰胺键连接在它的氨基上,磷酸以酯键连接在它的1-羟基上。鞘磷脂也是亲水脂两性分子,是高等动物神经组织中含量最丰富的鞘脂类(鞘氨醇是鞘磷脂的主要成分,故亦属于鞘脂类)。磷脂能在生物体内合成并快速地周转。
结构及命名
磷酸甘油酯甘油分子的中央碳原子是不对称的。天然的磷酸甘油酯都具有相同的立体化学构型,属于L系(见图)。
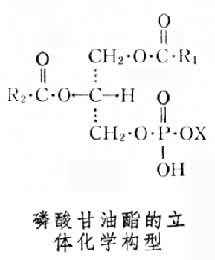
根据IUPAC-IUB国际委员会制定的脂质命名原则,磷酸甘油酯中:如X为胆碱,则应命名为:1,2-二酰基-sn-甘油-3-磷酰胆碱,亦称L-3-磷脂胆碱,俗名卵磷脂。图上构型中R1,R2代表脂肪酸链,X为连接在磷酸上的小分子化合物;名称中sn为立体化学专一编号。
磷酸甘油酯分子内部既含有强极性基团同时也含有强非极性基团。两个脂肪酸链形成非极性尾,而含磷酸的一端是极性头部。各种磷酸甘油酯的差别主要在于其极性头的大小、形状和电荷的差异。L-磷脂酸是最简单的磷酸甘油酯,磷酸基团上不连接任何小分子化合物。它是各种磷酸甘油酯的母体化合物,广泛地存在于细胞内,但仅有痕量,因为周转率很快,是合成各种磷脂和脂肪的关键中间产物。
每一种磷酸甘油酯都不是单纯的化合物,如磷脂酰胆碱分子内脂肪酸组成就是多种多样的。绝大多数磷酸甘油酯C-1位上以饱和脂肪酸为主,而C-2位上不饱和脂肪酸居多。
磷酸甘油酯分子中的碳氢链并不是无例外地以酯键连接在甘油的羟基上。缩醛磷脂的甘油分子中第一个碳原子由顺式烯醚键连接碳氢链,第二个碳原子以酯键连接长链脂肪酸。极性头通常是乙醇胺。另外还有一种醚磷脂是缩醛磷脂的还原产物,甘油分子的C-1以醚的结构连接碳氢链,这种化合物比较罕见。
鞘磷脂鞘磷脂与磷酸甘油酯的差别在于脂肪酸残基是连接在鞘氨醇的氨基上,“X”基团是通过磷酸连接到鞘氨醇的C-1羟基。“X”通常为胆碱或乙醇胺。鞘磷脂分子内的鞘氨醇碳链和脂肪酸碳链形成非极性尾,含“X”的磷酸端为极性头,也是亲水脂两性分子。神经组织鞘磷脂内的脂肪酸限于硬脂酸、廿四烷酸和神经酸。脾脏和肺脏鞘磷脂内的脂肪酸主要是棕榈酸和廿四烷酸。长链鞘氨醇有两类:鞘氨醇型和4-羟基双氢鞘氨醇型(亦称植物鞘氨醇型)。各种不同的鞘氨醇的差别在于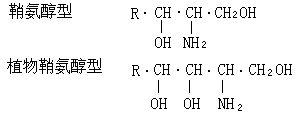 碳链长短(C14~C24);双键数目与构型;碳链分支(异-和反异-)生物体含有各种不同的长链鞘氨醇,在高等动物中,依进化趋势其碳链加长,不饱和度增加;植物和真菌的长链鞘氨醇含有三个羟基;海洋无脊椎动物以双不饱和化合物为主(表1)。
碳链长短(C14~C24);双键数目与构型;碳链分支(异-和反异-)生物体含有各种不同的长链鞘氨醇,在高等动物中,依进化趋势其碳链加长,不饱和度增加;植物和真菌的长链鞘氨醇含有三个羟基;海洋无脊椎动物以双不饱和化合物为主(表1)。

磷酸甘油酯的性质及类别
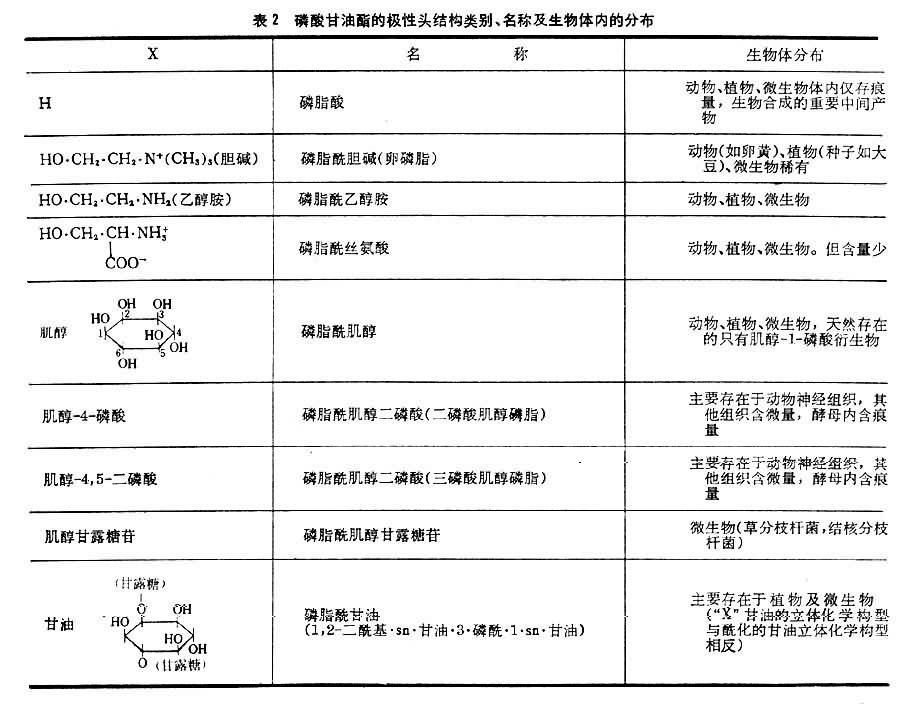
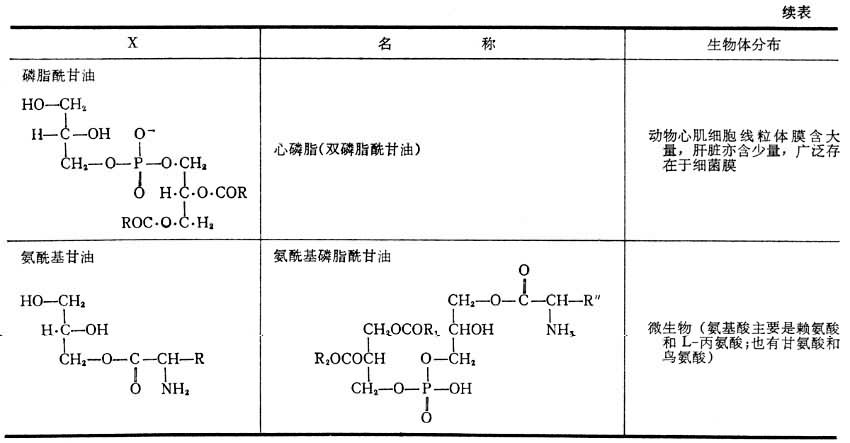
纯的磷酸甘油酯是白色蜡状固体,暴露在空气中变为棕红或棕黑色,这是因为其中的不饱和脂肪酸被氧化形成过氧化物的聚合物之故。磷酸甘油酯能溶于含少量水的非极性溶剂;与水相混合自动形成微团或片状双层结构,亲水的极性头面向水相,而疏水尾则互相聚集于微团内侧。这种脂质(磷脂和少量糖脂及胆固醇)双层是生物膜的基本结构。各种磷酸甘油酯所含极性头的大小,形状和电荷都不一样。磷脂酰胆碱,磷脂酰乙醇胺是既带有正电荷又带有负电荷的两性离子;磷脂酰丝氨酸分子在pH=7时带有两个负电荷和一个正电荷。这三者结构相似,亲水性强。具有强亲水极性头和疏水尾结构的化合物都有优良的去垢作用。表2列出磷酸甘油酯的极性头结构类别,名称及在生物体的分布。缩醛磷脂在肌肉和神经细胞膜中含量丰富。醚磷脂比较罕见,存在于某些种动物的红细胞及蛞蝓体内。脑磷脂是一个旧的俗称,指由磷脂酰乙醇胺和磷脂酰丝氨酸以及少量其他脂质组成的混合物。
磷脂的生物合成
磷脂酸是合成磷脂的前体,同时也是合成三酰甘油的前体。有几条不同的途径生成磷脂酸。
磷脂酸经两条途径合成不同的磷脂,两者都利用胞苷三磷酸辅因子。其一是磷脂酸经磷脂酸磷酸酯酶水解产生二酰甘油,后者与胞苷二磷酸胆碱或胞苷二磷酸乙醇胺作用产生磷脂酰胆碱或磷脂酰乙醇胺。在这里,胞苷二磷酸是“碱基” (X)的载体。全程合成始于胆碱或乙醇胺的磷酸化,活化的胆碱或乙醇胺与胞苷三磷酸作用形成CDP-胆碱或CDP-乙醇胺。
(1)甘油磷酸途径
(2)脂酰基磷酸二羟丙酮途径
(3)甘油二酯激酶途径
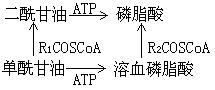
另一途径是磷脂酸与胞苷三磷酸反应生成“活化的磷脂酸”即胞苷二磷酸二酰甘油,后者能与丝氨酸或肌醇作用分别合成磷脂酰丝氨酸(细菌合成途径)或磷脂酰肌醇。磷脂酰肌醇经磷酸化反应能产生二磷酸肌醇磷脂和三磷酸肌醇磷脂(动物脑)。胞苷二磷酸二酰甘油还能与sn-甘油-3-磷酸反应产生3-sn-磷脂酰-1'-sn-甘油-3'-磷酸,再经水解切除磷酸基团从而合成磷脂酰甘油。后者再与CDP-二酰甘油作用就合成了心磷脂(动物体)。细菌利用2分子磷脂酰甘油缩合合成心磷脂。
动物体合成磷脂酰丝氨酸是通过酶促交换极性头的反应。动物体也能由磷脂酰乙醇胺的直接甲基化合成磷脂酰胆碱。磷脂酰丝氨酸经脱羧又能转变成磷脂酰乙醇胺。极性头的交换反应和磷脂之间的相互转变在改变膜磷脂的极性头分子结构上起重要作用。
鞘氨醇的氨基被长链脂肪酸酰化产生的N-脂酰鞘氨醇(神经酰胺)与CDP胆碱作用合成鞘磷脂。
磷脂的酶促降解及生理功能
磷酸甘油酯在生物体内经磷脂酶水解成为甘油、脂肪酸、磷酸和各种小分子化合物(X)。磷脂酶A1,A2,B,C,D分别作用于磷酸甘油酯分子内的不同酯键。
磷酸甘油酯的降解代谢通常不一定进行到底,部分降解的中间产物还能被细胞重新利用。对细胞来说,重新利用中间产物一方面节省能量,不必一切都从头合成;另一方面也能按照机体的需要改制磷脂分子。
催化磷脂降解的各种酶在磷脂周转和改造中起重要作用。如磷脂酶 A(包括A1和A2)能催化产生小量溶血磷脂,它们和酰基转移酶协同工作,对机体脂质内脂肪酸组成的调节改组起重要作用,是机体适应细胞需要“定做”具有特定脂肪酸成分的脂质的主要方式之一。磷脂酶A2能从膜磷脂释放花生四烯酸作为“在位”合成前列腺素的前体。磷脂酶A催化产生溶血磷脂富集的膜区,从而改变膜的通透性屏障。磷脂酶和酰基转移酶还能修复和更新那些由于自身氧化或其他损伤而造成的劣质磷脂分子,维持膜磷脂的完整。