[拼音]:heci gongzhen
[外文]:nuclear magnetic resonance, NMR
是磁矩不为零的原子核,在外磁场作用下自旋能级发生蔡曼分裂,共振吸收某一定频率的射频辐射的物理过程。核磁共振波谱学是光谱学的一个分支,其共振频率在射频波段,相应的跃迁是核自旋在核蔡曼能级上的跃迁。
基本原理
自旋量子数I不为零的核与外磁场 H0相互作用,使核能级发生2I+1重分裂,此为蔡曼分裂。核自旋在蔡曼能级之间跃迁的选择定则为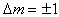 ,m为核磁量子数。可发生核自旋跃迁的两能级的能量之差
,m为核磁量子数。可发生核自旋跃迁的两能级的能量之差
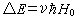
其中ν为旋磁比,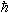 =h/2π,h为普朗克常数。
=h/2π,h为普朗克常数。
如果在上述静磁场H0存在的同时再加上一个方向与之垂直,强度远小于H0的射频交变磁场H 1,并且其频率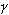 满足如下条件:
满足如下条件:
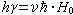
则原子核会吸收射频场能量,在两蔡曼能级之间发生跃迁,此现象为核磁共振现象。共振圆频率:
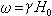
核磁共振频率因核而异,对于同一种核,共振频率与静磁场H0成正比。
主要参数
化学位移
同一种核在分子中因所处的化学环境不同,使共振频率发生位移的现象。
化学位移产生的原因是分子中运动的电子在外磁场下对核产生的磁屏蔽。屏蔽作用的大小可用屏蔽因子σ来表示。一般来说屏蔽因子σ 是一个二阶张量,只有在液体中由于分子的快速翻滚,化学位移的各向异性被平均,屏蔽因子才表现为一常量。
核磁共振的共振频率:
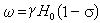
实际测定中化学位移是以某一参考物的谱线为标准,其他谱线都与它比较,即以一无因次的量δ表示化学位移的大小。常用参考物是四甲基硅(TMS)。

H参考,H样品分别是使参考物和被测样品共振的磁场强度,H0是仪器工作的磁场强度。
v参考,v样品分别是参考物和被测样品的共振频率,ν0是仪器的工作频率,化学位移的单位是(ppm百万分之一)。
由图1可见,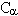 H,CβH及芳环上的质子的化学位移各异。其他核 (如13C,19F,31P等)的核磁吸收峰也因基团各异而有不同的化学位移值。即使同类残基的同一基团,因生物分子的特定构象而有不同化学环境,亦可有不同化学位移值。基于这点,人们可逐一识别各谱峰代表什么基团,因此NMR可广泛用于分子生物学,生物物理学以至生物医学研究。
H,CβH及芳环上的质子的化学位移各异。其他核 (如13C,19F,31P等)的核磁吸收峰也因基团各异而有不同的化学位移值。即使同类残基的同一基团,因生物分子的特定构象而有不同化学环境,亦可有不同化学位移值。基于这点,人们可逐一识别各谱峰代表什么基团,因此NMR可广泛用于分子生物学,生物物理学以至生物医学研究。
化学位移的大小受邻近基团的电负性、磁各向异性、芳环环流、溶剂、pH值、氢键等许多因素的影响。其中有3种效应常被用于生物学研究。
(1)环流效应:生物分子中常有含大π共轭电子云的芳环或芳杂环,如Phe、His、Tyr、Trp、嘌呤、嘧啶以及卟啉环。原子核相对于这些环的距离,方位不同,受大π电子云产生的附加磁场的影响不同,对各核化学位移的影响亦不同。环流效应常用于生物分子的溶液构象研究。
(2)顺磁效应:弤、Fe2+(高自旋态)、CO2+、Mn2+ 等顺磁离子及有机自由基(自旋标记化合物)中的不成对电子对周围核的化学位移及弛豫过程会有很大的影响,利用这个效应可研究顺磁离子周围基团的状况。
(3)pH滴定效应:在不同pH条件下,各解离基团的解离状况不一,造成附近基团有不同的化学环境,从而使得化学位移随pH变化。
耦合常数
核与核之间以价电子为媒介相互耦合引起谱线分裂的现象称为自旋裂分。由于自旋裂分形成的多重峰中相邻两峰之间的距离被称为自旋-自旋耦合常数,用J表示。耦合常数用来表征两核之间耦合作用的大小,具有频率的因次,单位是赫兹。
一般来说由于自旋耦合使高分辨核磁共振波谱变得十分复杂,但是当化学位移之差Δγ远大于耦合常数时,一个含有n个自旋量子数为I的核的基团将会使其邻近基团中核的吸收峰分裂为2nI+1重峰,并且这 2nI+1重峰的强度分布服从二项式系数分配公式(1+x)n。此为一级分裂波谱。
图1中各峰由于自旋耦合而产生谱线裂分。耦合常数的大小与外加磁场的大小无关,与分子结构有关即与两核之间键的数目及电子云的分布有关。一般来说,两核之间相隔 3个以上的化学键之间的耦合被称为远程耦合,J值很小。如果两核之间相隔四个或四个以上的单键,J值基本上等于零。
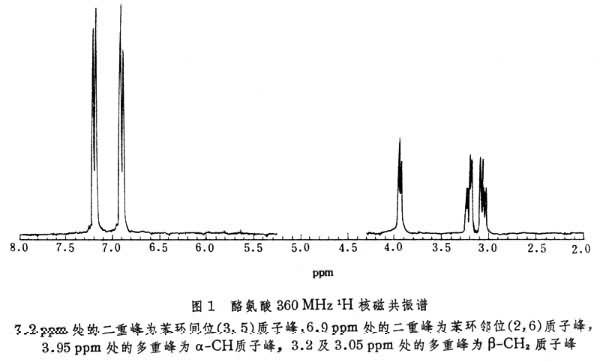
耦合常数可提供分子中二面角的信息。多肽和蛋白质中NH-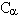 H耦合常数和围绕 -N 键的二面角
H耦合常数和围绕 -N 键的二面角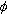 之间有如下的Karplus关系式
之间有如下的Karplus关系式
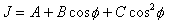
核苷酸的某些构象信息,亦可按一定公式由耦合常数测知。
谱峰强度
吸收峰下的面积,即吸收峰积分曲线的高度与产生该吸收峰基团的粒子数成正比。图 1中苯环间位质子峰,苯环邻位质子峰,α-CH质子峰,β-C啹质子峰的积分强度之比为2∶2∶1∶2。
弛豫参数
(1)自旋-晶格弛豫时间(纵向弛豫时间)T1,核系统与周围晶格相互作用,交换能量,使核系统恢复平衡,这一过程被称为自旋-晶格弛豫过程,自旋-晶格弛豫过程的快慢可用自旋-晶格弛豫时间T1来表征。T1的单位是秒。
(2)自旋-自旋弛豫时间(横向弛豫时间)T2,等同核之间的磁相作用被称为自旋-自旋相互作用。等同核之间相互交换自旋态并不改变系统的总能量,却缩短了系统在激发态的能级寿命。自旋-自旋弛豫时间T2是核处于激发态的能级寿命,以秒为单位,它与谱线宽度有关。
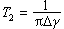
Δγ为谱峰半高宽(Hz)。 ③核奥弗豪泽效应。当一个强的射频场加到一组核上,使其中一个或多个跃迁被饱和,这时在空间相邻近的另一组核的共振信号的积分强度会因此而改变,这一现象被称为核奥弗豪泽效应,最早由A.W.奥弗豪泽发现。
核奥弗豪泽效应的大小可用NOE来表示:
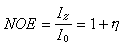
I0是未照射时信号强度,Iz是邻近基团被照射时的信号强度,η被称为核奥弗豪泽效应增强因子。
T1、T2、NOE都与分子运动的相关时间τ0有关,从中可以得到生物大分子内部动力学的信息,NOE还与两核之间距离的6次方成反比。
核磁共振谱仪
分为两大类,即连续谱核磁共振谱仪及脉冲傅立叶变换核磁共振谱仪。前者将单一频率的射频场连续加在核系统上,得到的是频率域上的吸收信号和色散信号。后者将短而强的等距脉冲所调制的射频信号加到核系统上,使不同共振频率的许多核同时得到激发,得到的是时间域上的自由感应衰减信号(FID)的相干图,再经过计算机进行快速傅立叶变换后才得到频率域上的信号。
历史发展
1946年美国斯坦福F.布洛克和美国哈佛大学E.M.珀塞耳小组几乎同时记录到液体样品和固体样品中的核磁共振信号。核磁共振波谱技术用于生物学问题始于50年代中,首次成功地用核磁共振波谱直接观察到生物大分子的是桑德斯和J.G.柯克伍德,他们于1957年报告了40MHz的核糖核酸酶的1H核磁共振波谱。同年O.雅德茨基和C.D.雅德茨基也证明核磁共振波谱可以精确地反映蛋白质的氨基酸组成。到目前为止,生物体中蛋白质、核酸、磷脂、多糖及其相应组分均可用 NMR技术研究,核磁共振波谱已成为在溶液态研究生物高分子空间结构与功能的最重要物理手段。
在生物学中的应用
核磁共振波谱技术用来研究生物大分子有如下特点:
(1)不破坏生物高分子结构(包括空间结构)。
(2)在溶液中测定符合生物体的常态,也可测定固体样品,比较晶态和溶液态的构象异同。
(3)不仅可用来研究构象而且可用来研究构象变化即动力学过程。
(4)可以提供分子中个别基团的信息,对于比较小的多肽和蛋白质已可通过二维NMR获得全部三维结构信息。
(5)可用来研究活细胞和活组织。
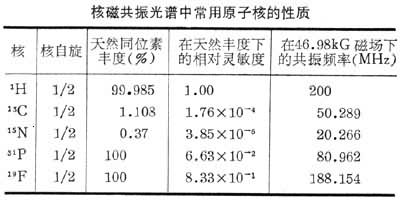
在生物学研究中最常用的是1H,13C,31P谱,此外还有15N,19F等。1H谱发展最早,1H在生物体中无所不在,核磁相对灵敏度高,故应用最广,包括用于核磁成像。缺点是含氢基团极多,谱线易重叠而不易解析。碳亦为生物体内重要元素,但12C自旋为零,13C天然丰度低,仅为1.1%,对等数量的核在相同磁场下其灵敏度只及1H的1.6%。其优点是化学位移范围宽,在宽带去耦条件下进行实验,波谱简单,易分辨,随着测定技术及13C标记方法的发展,13C 谱已有极广泛的应用。31P谱常用于活组织测定,观察ATP等含磷化合物的代谢过程,并已用于核磁成像。
NMR在生物学研究中范围很广。主要有:
分析研究
如确定生物分子成分及浓度,特别是可不破坏组织细胞而测知其中组分;确定异构体比例;确定分子解离状态;确定金属离子或配基是否处于结合状态;以及测定细胞膜内外的pH差异。
热力学研究
如测定酶与底物、配基、抑制剂的结合常数;测定可解离基团的pK值,特别是能测定生物大分子中分处不同微环境的同类残基的同类基团的不同pK值,这是其他方法所不及的;还可测定相变温度,ΔG等其他热力学参数。
动力学研究
如监测反应进程,测定各组分随时间的变化;通过变温实验和线形分析,测平衡过程的动力学常数,包括某些生化反应的反应速率,研究分子间(如酶与抑制剂,DNA与药物)相互作用的动力学过程。
分子运动研究
弛豫参数 (T1,T2,NOE)可用来研究生物高分子的动力学,以及生物膜的流动性。
分子构象及构象变化研究
目前用二维核磁共振技术加上计算机模拟已能独立确定小的蛋白质分子及核苷酸片段在溶液中的三维空间结构。改变物理化学因素或加入可与生物分子相互作用的其他物质,将会使核磁图谱发生变化,从而可用来研究这种构象变化。
活体研究
用31P,13C,1H磁共振方法测定活细胞,活组织以致整体的代谢物浓度及变化,测定细胞内pH值,观察药物或不同生理状况对代谢的影响。研究对象有微生物、植物、各种动物以至人体器官等。
二维核磁共振谱
自70年代末,特别是80年代以来二维核磁共振(2D-NMR)及核磁成像技术(NMRimage)迅速发展,已在生物学、医学研究中发挥越来越大的作用。
二维核磁共振的思想是1971年提出的,1976年R.R.恩斯特用密度矩阵方法对二维核磁共振实验进行了详细的理论阐述,自此二维核磁共振技术得到了非常迅猛的发展。1977年K.维特里希首先将此方法用于生物高分子,研究氨基酸和牛胰胰蛋白酶抑制剂,在此基础上发展了用二维核磁共振对蛋白质1H-NMR的单个谱峰全部识别的方法。
二维核磁共振谱S(ω1,ω2)被定义为二个独立的频率变量ω1和ω2的函数,它是由对时间域上的函数S(t1,t2)进行二维傅立叶变换得到的。
其基本原理如下:所有二维核磁共振波谱在时间域上都可分为四个时期,即准备期、发展期、混和期(可以没有)及检测期,其中发展期的时间间隔为t1,检测期的时间间隔为t2。
准备期一般比较长,自旋系统在这一时期达到热平衡。发展期加上一个或多个射频脉冲使核系统演化。混和期包括脉冲和延迟时间。在检测期,信号作为t2的函数被检测,并以发展期的时间间隔t1为参数。只改变发展期的时间间隔t1,重复多次实验,即可得到时间域的二维信号S(t1,t2),对此作二维傅立叶变换即得频率域的二维核磁共振谱S(ω1,ω2)。二维核磁共振谱可分为3类:
化学位移相关谱
这类实验可以将相互耦合的核或者具有化学交换的核,或是有交互弛豫的核的化学位移相关联。
J-谱
在J(耦合分解)谱中,决定共振峰位置的两个参数,即标量耦合和化学位移得到分开。
多量子谱
这类实验可用来检测多量子跃迁。K.维特里希将2D-NMR用于蛋白质结构研究,使得人们有可能对小的或中等大小的蛋白质,无需晶体结构资料,只根据一级结构序列和一套 NMR数据就可以推断该蛋白质在溶液中的构象。
对肽链中各基团的质子而言,存在两种相互关系,一种是通过化学键的J耦合关系,一种是通过空间的NOE关系(图2 )。蛋白质中上述两种相互关系可以通过二类2D-NMR实验找出:
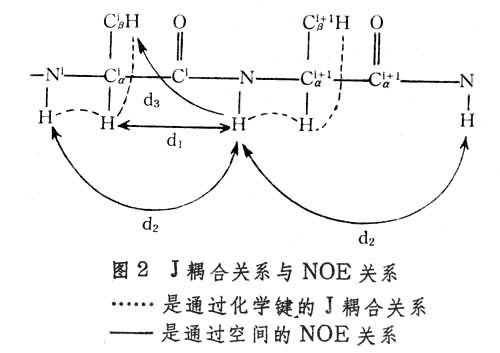
(1)二维相关谱(COSY) 如图3所示,对角线上的峰是一维谱峰,对角线以外的峰为交叉峰,交叉峰相对于对角线为轴对称,从一个交叉峰出发分别作垂直线与水平线与对角线相交,相交处的两峰所对应的核之间有 J耦合。因此交叉峰将标量耦合的质子相关联。
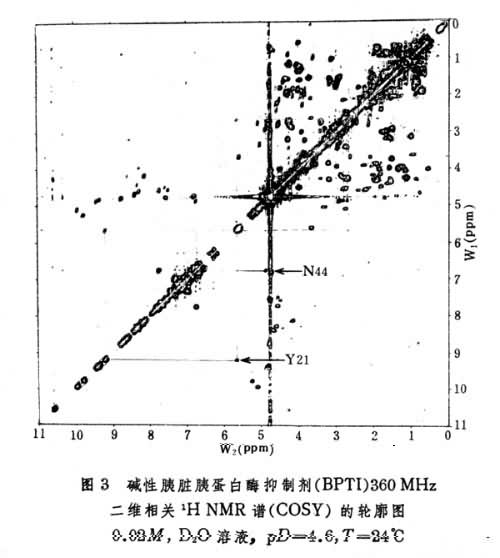
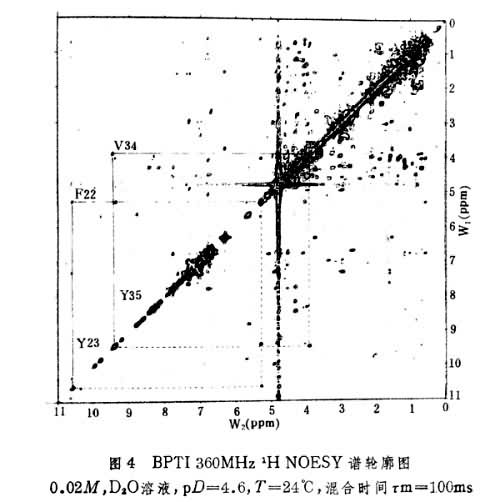
(2)二维核奥弗豪泽增强谱(NOESY) 如图4所示,其交叉峰将空间上比较接近,相互之间有NOE的峰相关联,这包括同一氨基酸残基中由共价键相连的质子如 CαH,CβH,CγH,在氨基酸序列中相邻残基之间主链上的质子,如C H和Ni+11H,以及在一级结构序列中分开较远而在三维空间结构上接近的质子。实际工作中将半张COSY谱,半张NOESY谱按对角线并接成图即可沿肽链按残基序列逐一识别出共振峰。此外NOESY谱还可提供1H和1H之间空间距离的信息,从而得到有关蛋白质二级结构的信息。
H和Ni+11H,以及在一级结构序列中分开较远而在三维空间结构上接近的质子。实际工作中将半张COSY谱,半张NOESY谱按对角线并接成图即可沿肽链按残基序列逐一识别出共振峰。此外NOESY谱还可提供1H和1H之间空间距离的信息,从而得到有关蛋白质二级结构的信息。
2D-NMR
目前已不仅被用来研究蛋白质,而且用来研究核酸以及蛋白质和核酸的相互作用。
核磁成像技术(NMR image)
与X射线断层成像(CT)相似,但其物理过程则完全不同。CT成像靠不同组织对X射线的吸收差异而成像;核磁成像则是因不同组织的自旋密度(1H或31P)或弛豫参数(T1或T2)不同而成像。核磁成像技术的基本原理是根据非均匀磁场造成的谱线的非均匀加宽中包含有核自旋空间位置和它们的分子环境的信息。若在静磁场H 0上再叠加一个线性磁场梯度G,则只有位于垂直于G 的同一平面中的质子才受到相同的磁场,即具有相同的共振频率,其强度正比于核平面中核自旋数,不同平面中质子共振频率不同,强度也不同,从而使谱线非均匀地加宽,这种加宽了的共振峰实际上就是样品中核自旋密度在G 方向上的投影。改变磁场梯度的方向即可得一系列共振吸收谱,用计算机进行图象重构,则可得到被测物体在某一平面上的投影图。除上述投影重建法外,近年来还发展了敏感点扫描法,选择性激发法及场聚焦法等成象方法。NMR 图象除了给出质子或其他一些核的自旋密度的空间信息外还能得到弛豫时间的空间信息。目前核磁成像已用于临床诊断,其优点是对软组织敏感,无X射线损伤,不仅能显示形态结构,还可提供不同组织的生理、化学特性,可以自由选择剖面。
- 参考书目
- O.Jardetzky,N M R in Molecular Biology, Academic Press,New York,1981.
- K.Wüthrich,N M R in Biological Research Peptides and Proteins, North Holland Amsterdam, Biomedical Press,1976.
- L.J.Berliner,J.Reuben eds,Biological Magnetic Resonance, Vo1.4,Plenum Press, New York,1980.