[拼音]:haiyang shengtai
[外文]:marine ecology
海洋生物之间及海洋生物与其海洋环境之间的相互关系。海洋是生命的发源地,其中孕育着种类繁多的海洋生物,每年为人类提供大量资源。
海洋环境
地球上海洋面积共3.6亿平方公里,约占地球总面积的71%,为陆地面积的2.4倍。海洋之间并无隔断,由于海水的水平和垂直运动,还不断地进行着水体和物质的混合和交换。海洋平均深度约3820米。目前已知海洋最深处超过11000米。通常水深每增加10米,就增加1个大气压。因此海底最深处的压力可达1100个大气压。
海底地形
海底构造复杂,有海底山脉、高原、盆地、峡谷和海沟等各种地形。通常将海域分为:
(1)大陆架,海洋靠近陆地的部分,从沿岸起缓慢往外倾斜,平均坡度仅0°07′,即向外延伸1公里,平均加深1.5米左右。大陆架的宽度与陆地地形有关,有些海洋几乎没有大陆架,而中国沿海大陆架宽度有几百公里。大陆架水深可达200米。
(2)大陆坡,在大陆架以外,其主要特点是坡度明显增大(有的可达20°~30°),水深也急剧增加,平均可达2440米左右。
(3)大洋盆地,在大陆坡以外,地形广阔而平坦,是海洋的主要部分,水深可达6000米左右。
(4)海沟,狭而长,水深超过6000米以上,最深的海沟是马里亚纳海沟,水深达11022米。
海洋的水平分区
与大陆架相对应的浅海称为近岸区或近海区,大陆坡以外的广大海域称为大洋区。
近海区面积小,仅占海洋总面积的8%,但由于陆地大量营养物质注入,因此是生物生长、繁殖的重要场所。
海洋的垂直分层
大洋区的水层可垂直划分为若干带。
(1)上层带,下限为浮游植物的补偿深度,即光合作用产生的氧恰与呼吸作用消耗的氧相等的深度,其具体数值取决于光强和水的透明度,一般10~120米,平均约为50米。
(2)中层带,起于上层带的下限,下至200~300米深处。
(3)次中层带,水深可达600~700米。
(4)半深带,水深由600~700米至2000~2500米,几乎无光。
(5)深层带,水深2500~6000米,无光。
(6)深渊带,水深在6000米以下,无光。
海洋水温通常由表层向下递减,到达一定深度,温度降低显著,即温跃层。深层带和深渊带水温一般是1~4℃。许多海洋生物往往只能在一定水层生活,如浮游植物生活在上层带。
海洋纬度自然带
海洋表层水温在一年中的变化,随水体所处纬度而定。按照表层水温可把海洋划分为若干自然带。
(1)热带,表层水温最低在20℃以上,温度变化范围小,一般仅2~3℃。
(2)亚热带,最低水温在15℃以上,温度变化范围较大,可达6~7℃以上。
(3)温带,水温在15℃以下,可近于0℃,温度变化范围最大。
(4)近极区,水温最高不超过10℃,温度变化范围不超过2~3℃。
(5)极区,水温最高不超过0℃(或5℃),温度变化范围很小,沿岸常有冰层覆盖。
海流
对海洋环境起着积极主导的作用。海流包括潮流、风海流、坡度流、升降流(上升流和下降流)、密度流、涡流和大洋环流等。大洋环流是大洋海流的骨干,而上升流区域则是重要的渔场。
太平洋黑潮暖流经中国台湾海峡的东面,沿东海边缘向东北方向流去。它的两支分流对中国沿海有明显的影响。一支分流经巴士海峡流入南海;另一支分流经东海注入黄海,在夏季甚至可达渤海。这两支海流的方向大体都是由南向北,是暖性的。中国沿海另一支海流是中国沿岸流,北起黄海和渤海,沿海岸南下,经台湾海峡注入南海。沿岸流受大陆气候影响,冬夏温差大,沿途容纳了大量淡水,所以盐度也较低。在台湾以东海域、舟山附近、海南岛东北部、粤东和闽浙沿海有上升流。除海流外,波浪、潮汐、光、温度和水压力等都是影响海洋生物的重要物理因素。
海水的化学组成
已发现海水含有80多种元素,其化合物的种类则更多。这些物质以离子态(阳离子、阴离子、络合离子)、悬浮态、胶体以及气泡等形式存在于海水中,海水总的含盐量在35‰左右,但在河口可低至1~2‰。个别水蒸发量大的海域含盐量偏高,如红海为40‰。按组分含量的不同,可把海水中的化学成分分为5类:
常量元素
如钠、镁、钙、钾、锶、氯、硫、碳、溴、氟、硼等,占海水中溶解盐类的99.8~99.9%,它们在海水中的含量基本不变。
营养元素
包括氮(以硝酸盐、亚硝酸盐和铵等形式存在)、硅、磷、铁、锰、铜等,这些元素与生物的生长关系很密切。
微量元素
种类很多,但总量却非常少,仅占总盐量的0.1%左右,如锂、铷、铯、锌、铀等。
溶解气体
主要来自海气交换,如氮、氧、氩、二氧化碳、氖、氦等,在缺氧情况下,还会出现硫化氢、甲烷等。
有机物
包括碳水化合物、脂肪、蛋白质和元素有机化合物等等。部分来源于大陆河流输入,大部分来自海洋生物的分泌、排泄和生物死体的分解。
海水的盐度、pH值、含氧量、二氧化碳含量、营养盐和有机物以及一些微量元素的含量,对海洋生物都有直接的影响。
海底沉积
海底的地形、结构以及沉积物的组成、理化特性(如粒度大小、有机质含量、氧化还原电位等)是影响底栖生物生活的重要环境元素。海底沉积物来源大致是:陆源性沉积、陆上或海底火山的喷出物、无机性沉淀或宇宙产物,以及由生物死体所造成的细泥。在浅海区域,底部沉积物的组成主要受陆地物质来源的影响,海流对沉积物的转移起主导作用。绝大部分深海或大洋底层的沉积物,都是由海洋生物的壳和骨骼所造成,在大洋的底部,生物尸体的沉积有的竟厚达数千米以上。在近海区,沉积速率一般较高;而在大洋区,沉积速率很慢,每千年仅沉积1厘米左右。
海洋生物
海洋中几乎到处都有生物,但不同的环境,生物群落的种类组成和结构,以及各种群数量、个体大小、形态、生理生化特性等都很不同。海洋生物分布的格局是与海洋环境相互作用、协调进化的。一般常按生活方式将海洋生物分为浮游生物、游泳动物和底栖生物3类。
浮游生物
无游泳能力或游泳能力弱,悬浮于水中随水流移动,包括细菌、浮游植物(如硅藻、甲藻等)和浮游动物(如水母、腹足纲软体动物的翼足类、异足类,许多海洋动物的幼虫等)。多数终生营浮游生活,称为永久性浮游生物;少数种类仅于生活史的某个阶段营浮游生活,称为阶段性浮游生物,如许多海洋动物的幼虫;也有些原非浮游生物,被水流冲荡而出现在浮游生物中,如某些低等甲壳类的介形类、涟虫类等,称为暂时性浮游生物。浮游植物只能生活在有光的水层;浮游动物则不然,有的可以生活在千米以下的深水中,而且多数能在水中作垂直移动。浮游生物一般体重轻(外壳重量轻、体内脂肪含量高,富有粘液)、沉降阻力大(身体相对面积大,体表多刺毛、突起,群体连成片)或者具有纤毛、鞭毛而有一定的运动能力。
游泳生物
指那些具有发达的运动器官而游泳能力强的动物。包括鱼类、哺乳动物(如鲸、海豚、海豹)、爬行动物(如海蛇、海龟)、软体动物(如乌贼、章鱼)和一些大型虾类(如对虾、龙虾)等,它们的游泳能力、速度和方式也有很大差异。有些种类能横跨大洋作长距离的洄游,如金枪鱼等;有的游泳很快,如剑鱼每小时速度达70公里以上。海鳗以整个身体弯曲摆动向前游动,乌贼和章鱼则从漏斗口向外喷射水流以推动身体反向运动。它们有时生活在上层水域(如太平洋鲱鱼),有的生活在中层或底层水域(如小黄鱼、真鲷、牙鲆)。游泳动物除了有发达的游泳器官外,身体大多呈流线型,以减小阻力,提高游泳速度。
底栖生物
生活在海洋水域底部和不能长时间在水中游动的各种生物,包括底栖植物(几乎全部大型藻类和红树等种子植物),底栖动物(海绵、腔肠、环节、线形、软体、甲壳、棘皮、脊椎等门类均有底栖种)。底栖生物按其与底质的关系,又可区分为底上、底内和底游3大生活类型;在岸边还存在潮间带生物。
底上生物
生活于硬质海底。包括在各种底质上营固着生活(如海葵、水螅、藤壶、牡蛎、柄海鞘)、附着生活(如紫贻贝、附着硅藻、大型藻类)以及匍匐海底(如马粪海胆、刺参和红螺等)的动物。固着生活的动物运动器官退化,但触觉器官发达,被动取食,幼体营浮游生活,靠海流扩大其分布区域。固着或附着在船底、浮标、工厂排污管道、海上石油平台和海上其他设施的生物被称为污损生物(包括微生物、藻类和动物)。它们对交通、军事和生产均有危害。
底内生物
生活于软质海底。有的分泌粘性物质形成管子或用分泌物粘结沉积颗粒成管子而栖身其中(如巢沙蚕、磷沙蚕等);有的具有发达的挖掘器官,将身体埋于泥沙中(如黄岛长吻柱头虫、中国蛤蜊、文昌鱼等);有的钻木(如船蛆)、凿石(如波纹沟海笋)。
底游生物
有一定的运动能力,能在水底缓慢地行动(如梭子蟹)。
潮间带生物
一类特殊类型的底栖生物。经过长期的进化,底栖生物对潮汐的涨落、海浪的冲击、阳光的暴晒、温度的变化等有很好的适应能力。由于潮间带生态位的差异,生物在潮间带的分布很有规则,群落结构比较清楚(图1)。
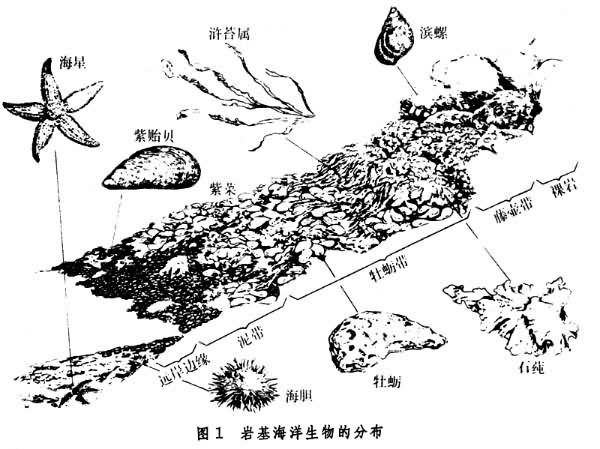
海洋生物的地理分布和群落结构,明显地受海洋理化环境的影响。温度对许多生物的地理分布是主要限制因子,例如,典型热带性珊瑚礁,只能在月平均水温不低于23.5℃的海域形成。中国沿海红树林的生长,最北达到北纬25℃(福建省南部)附近,冬季平均水温约15℃,再往北即不能生长。中国沿海的软体动物,有的仅生长在南海区域(如鹦鹉螺、虎斑宝贝以及大多数珊瑚种类);有的生长在南海和东海,而不生长在黄海和渤海(如蛇螺、台湾枪乌贼);有的仅生长在黄海和渤海(如盘大鲍、栉孔扇贝);有的在4个海域都能生长(如西施舌、大竹蛏)。
海流和水团决定了浮游生物群落的种类组成。根据对南黄海和东海浮游生物的调查,发现群落结构与这一海域海流和水团的性质密切相关,大致可分为:
(1)南黄海中部群落,分布区域与黄海冷水(低温、高盐)分布范围相吻合;
(2)沿海群落,分布在沿岸30海里以内范围,受中国沿岸流(广温、低盐)控制;
(3)东海外海群落,处于台湾暖流和黑潮暖流流经的区域(高温、高盐)。每个群落均有代表种,且一年四季又有所变动。
海底沉积物的特性对底栖生物群落有明显影响。据调查(1963),中国青岛胶州湾底栖动物的区系完全符合与地形、海底沉积及淤泥程度相依的规律,可分为5个明显的生物群落(表1)。
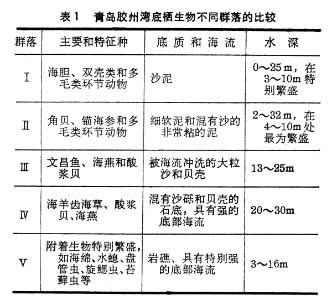
海洋生物大多数种类的种群大小呈现周期性波动,波动周期的长短不一。有些鱼类种群的波动周期性要经过几十年才表现出来。例如,挪威的鳕鱼的捕获量在将近50年中出现了两个高峰(图2)。有人注意到,鳕鱼的捕获量与松树年轮的宽度惊人地一致,而年轮的宽度是当年降雨量多少的反映。英吉利海峡的大西洋鲱鱼、波罗的海鲱和沙丁鱼的产量也呈现长周期的波动,且与表层水温和水中磷酸盐的含量的波动密切相关。又如,近百年来,太平洋鲱鱼在中国北方沿海的产量有很大的波动。据调查,黄海鲱鱼产量呈长周期变动,它在20世纪初、30年代末、60年代末和70年代初,在黄海北部曾有过3次丰产高峰。经分析,鲱鱼的大发生主要与亲鱼数量、亲鱼世代等内因密切相关,此外又与太阳活动引起的气象、海洋状况、海洋生物(饵料)和捕捞强度等外因密切相关。
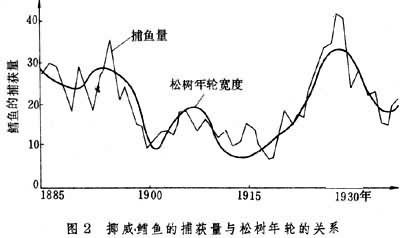
海洋生态系统
生态系统的最基本特征是,能量通过食物链不断地单向流动,物质在食物链与无机环境之间不断地反复循环。(图3)表示出海洋生态系统的结构。
海洋食物链
海洋生物群落各个种之间的食物关系十分复杂,可连结成链状或网状(食物链或食物网)。过去只注意海洋的牧食食物链,但20世纪60年代后发现,在河口、内湾、红树林和浅海水域,碎屑食物链也很重要。碎屑主要来自海洋植物的碎屑、动物没有消化的残渣、和陆地碎屑的注入等。已知有不少海洋动物以碎屑为食物,在河口及浅海水域,估计总的初级生产量大约有50%通过碎屑食物链。
食物链的长短不一,长的可有5~6个环节,杂食性动物还可同时占据不同的营养级。
海洋动物对食物的选择不如陆地动物严格,能摄食多种食物。在海洋中尚未发现在陆地上见到的那种单食性种类。例如,中国闽南-台湾浅滩渔场的二长棘鲷是底栖鱼,其食料生物种类广泛,包括长尾类(细鳌虾、褐虾)、底栖端足类(钩虾亚目)、瓣鳃类(短齿蛤、鸟蛤、帘蛤)、介形类(尖尾海萤)和多毛类(沙蚕、似柱虫),还摄食樱虾类(日本毛虾)、糠虾类、辐蛇尾、海胆类、星虫类、腹足类、掘足类、短尾类、海葵类和鱼类。海洋动物的捕食与被捕食关系相当复杂。大多数动物的幼体和成体的食性往往不同,可以处于不同的营养级。鲱鱼的幼体为箭虫所摄食,而箭虫却是成体鲱鱼的一种重要饵料。
生产者
海洋中,能行光合作用的生物有5类:大型定生藻、海洋被子植物、浮游植物、底栖硅藻和光合细菌。尽管在近岸海域大型定生藻的初级生产也很可观,但就整个海洋初级生产力来说,个体很小的浮游植物是主要生产者,这与淡水环境近似而与陆地环境有很大差别。小型浮游植物所制造的有机物约占海洋初级生产力的95%。因此,海洋初级生产力调查,主要是测定海洋浮游植物的光合作用速率。
影响海洋初级生产力的因素主要是光、海况、营养盐、浮游植物的种类、生物量和被捕食情况等。
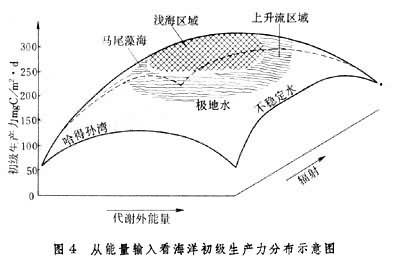
海洋生态系统的能量输入可分为两大类,一类来自太阳的辐射,另一类来自物理环境。运动引起的能量被称为代谢外能量。只有太阳辐射能高、代谢外能量输入也高的海域,才能够得到高的生产力,如近岸水域和上升流区域(图4)。
光是海洋初级生产者最重要的环境因素,由于海水对光的吸收和海水中悬浮物质对光的散射,光线进入水体后很快被衰减。在真光层下限,入射光强大约只相当于水表面光强的 1%。光透入水层的深度依海域所处的纬度、季节、海水的混浊度而变化。在清澈的大洋水中,透入水层的主要是430微米的光波,最深可达200米。在沿岸水域,由于短波长的光大部分被散射,进入水中的主要是550微米的光波,穿透深度小于大洋水。
光是海洋初级生产力的一个重要限制因素。若已知光随水深的衰减率、太阳辐射的量以及水中浮游植物色素的数量,即可按以下公式:
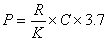
计算出浮游植物的生产力。式中的 P为浮游植物的光合作用率,即每天每平方米有机碳的克数;R为每天每平方米水表面所受到的太阳辐射能的相对光合作用率;K为消光系数(每米水深);C为在水柱中每立方米水体叶绿素的克数。
有人曾用此公式计算阿拉斯加湾的初级生产力,所得结果是1.34克(有机碳)/(米2·天),这与实测初级生产力1.50克(有机碳)/(米2·天)很接近。
浮游植物的生长离不开光,但并非所有的浮游植物都能适应强光。因此浮游植物的峰值大多出现在其光强相当于入射光能量大约 1/3的水深处。有些能够移动的浮游植物,如甲藻,还能根据光强调节所处水深。不同种类的浮游植物对光的利用率有差异,硅藻还借助辅助色素提高对光的利用率。
海洋初级生产力除了有地域差异时,还有日变化和季节变化。在北纬50°以北海域,初级生产力的高峰通常出现在中午。而在亚热带和热带海域,高峰出现在中午以前。青岛胶州湾浮游植物有明显的季节变化,在2月和8月,浮游藻细胞总量呈现出高峰,而6~7月和9月则是低谷期。
海流和水团对初级生产力有明显的影响。在西北太平洋亚北极水域和混合区水团的初级生产力高于亚热带水团和黑潮流域。
浮游植物种类的组成和比例是水域初级生产力高低的重要标志。硅藻占优势的水域,生产力也高。青岛胶州湾及附近水域,浮游植物种群结构以硅藻和甲藻两大类为主,尤其是硅藻,无论在种数或细胞个数,都占绝对优势。湾内硅藻的细胞数量可占浮游藻细胞总量的99.9(1979.2)~96.0%(1997.7)。
影响海洋初级生产力第二个重要因子是营养盐。其中氮和磷在浮游植物生长中起最主要限制作用,在某些水域束毛藻能直接从水中吸收分子氮,因而磷成为限制因子。相反,在某些沿岸水域,由于有大量含磷废水输入,氮又可能成为限制因子。一般情况下,氮磷比大约是15~16,比率在10~20范围内均能满足生长的需要。在温带浅海水域,春季浮游植物的大量繁殖,主要与冬季水混合时将大量营养盐输入真光带有关。上升流区域初级生产力高,主要是大量营养盐从海底带上升到水表层,使硅藻等浮游藻大量繁殖所致。根据营养盐含量和初级生产力高低,水域被分为富营养、寡营养和中等营养水域。
除了氮、磷外,硅是硅藻生长的一个重要因子,铁和一些溶于海水中的有机物,如螯合剂、维生素等,通过对浮游植物生长、繁殖的促进或抑制,也直接对初级生产力产生影响。
在沿海水域,大型海藻、底栖微型藻、红树林、海草等的生产力也值得重视。这是因为它们的生产量也相当可观。据估计西太平洋地区海藻的潜在年生产量可达3.0×106吨(鲜重),全球底栖微型藻可达109吨(碳)。不少海洋动物直接以它们为饵料,比如鲍鱼直接吃海带类海藻,有些虾类直接以红树林碎屑为饵料。
初级消费者
植食动物多是滤食性种类,以浮游植物或有机颗粒为食。主要包括桡足类等小型甲壳动物、被囊动物、毛颚动物、水母等浮游动物,贻贝、牡蛎、扇贝等底栖动物,以及植食性鱼类等。浮游动物与浮游植物杂居在同一水层,在水中能作垂直移动,其种群组成和数量也有明显的季节变化。
海洋动物的生产力,指一定时间内一群动物所增加的身体物质总重量。但浮游动物生产力测定有相当困难,常只能间接估算。
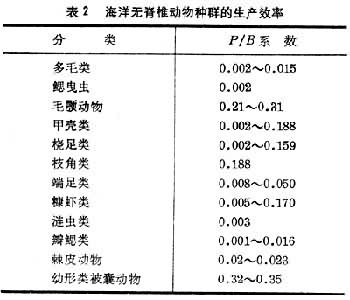
动物的生产效率,通常用P/B表示,其中P表示生产量,B表示某一种群的生物量。根据现有的资料,海洋无脊椎动物的生产效率大致如表2。
南极磷虾有很高的群聚性和生产量。曾观察到有一群磷虾连绵几平方公里,在水中厚达1800米,总计约有1000万吨。据估计南极海洋的磷虾现存量约达20亿吨。
次级生产力与个体大小、发育阶段、饵料质量和数量、摄食率、同化效率、温度等有直接关系。有的动物滤食率很高,比如一个海鳟类个体,每分钟可滤食100毫升,它对饵料没有选择性。海洋植食性动物的同化效率比陆地动物高,可达80~90%。
除摄食外,有些无脊椎动物以及海洋动物幼虫,还能直接从水环境中吸收溶解的有机物质(氨基酸、脂肪酸、多醣及其水解产物)。无脊椎动物幼虫从溶解有机物质直接吸收的碳可能占其代谢需要量的21~57%。曾有报道,海岛哲水蚤对无机和有机颗粒有识别能力,能从混合物中选择摄食有机物。
次级消费者
即肉食动物,以植食性动物或其他肉食性动物为饵料。因此,不同肉食性动物在食物链上可以处于不同的营养级。例如,北方河口的动物,依其营养关系可分为植食性动物、杂食性动物、低级肉食性动物、中级肉食性动物和高级肉食性动物 5类。据调查(1981),闽南-台湾浅滩渔场66种经济鱼类中,低级肉食性动物主要摄食植食性动物和杂食性动物,其种类和数量最多,共有42种,如金色小沙丁鱼、鲐鱼、二长棘鲷、银黄姑鱼、青石斑鱼、日本竹荚鱼等。中级肉食性动物主要摄食低级肉食性动物以及植食性和杂食性动物,其种类和数量比较少,共有16种,如大黄鱼、中国团扇鳐等。高级肉食性动物主要摄食低级和中级肉食性动物,以及杂食性动物,其种类和数量最少,只有8种,如带鱼、日本马鲛、路氏双髻鲨、沙拉真鲨等。
有些肉食性动物的摄食量很高。比如生活于南极海洋的蓝鲸,每餐可摄食1吨的磷虾。海洋动物的食性广泛,不仅在生长的不同时期采食种类不同,而且随着季节的不同,食物的组成也有差异。
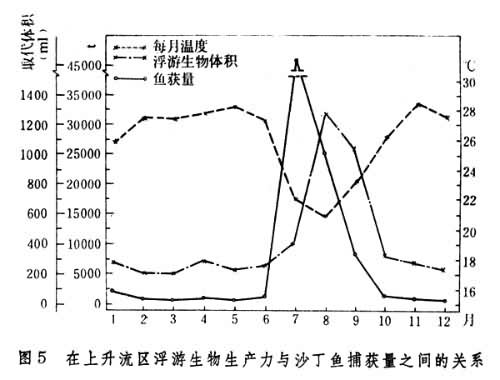
通常鱼获量与浮游生物的生物量呈正相关。在沿岸,每年7~9月有上升流,在此期间水域的浮游植物和浮游动物的生物量呈现峰值,而沙丁鱼也出现最大捕获量(图5)。
主动捕食者每日食量要高于消极等食的种类。比如,鲐鱼是主动掠食者,每天所获得的食物可达其体重10~25%;有些底栖鱼类主动性较差,每天摄食量仅1~3%。
海洋动物排出的粪便(粪粒、粪球)含有未消化的有机残渣,沉入海底后,成为某些底栖动物的饵料。
沿着海洋食物链营养级位而上,生物个体也逐渐增大。捕食者与捕获物的比例,按重量比是100:1,按长度比是4.6:1。鲸是个特例,它是现今地球上最大的动物(体重可达百吨),但却以个体仅几厘米的磷虾为饵料。
食物链的长短不一,以微型浮游植物为初级生产者的大洋水域,食物链长些,可达5~6个环节,如下左:
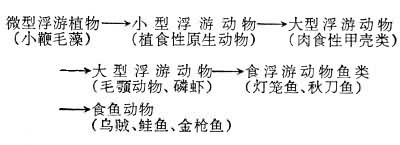
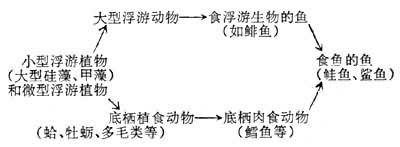

大陆架水域的食物链,主要以小型和微型浮游植物为初级生产者,食物链一般短于大洋水域,如上中:以大型浮游植物为主要初级生产者的上升水域的食物链大多很短,如上右:
分解者
主要是异养的微生物,它们借分解海洋动植物的死体和其他有机物质获得能量,同时把有机物逐渐降解还原为无机物。海洋中的碳循环、氮循环、磷循环等与陆地生物一样都离不开微生物的作用。同样地,海洋微生物对于净化有机物污染,如石油、有机农药等污染起积极的作用。水域的净化离不开微生物。
越来越多的事实表明,海洋细菌不仅起着还原者的作用,而且还是许多种海洋动物的直接饵料。海洋细菌,无论是在水中和海底沉积物里,其生物量都相当可观。据对大西洋一些浅水站位调查,细菌的生物量约占总的微小生物量的9.4%;而在大陆坡水中,细菌所占的比例增至52.5%。在某一个大洋站位,细菌的生物量在水中所占的比例高达94%。已查明,某些海洋浮游动物的食物来源中细菌所占的比例,可达30~50%。
能流
海洋生物的能量转换效率(生态效率)要比陆地生物的高,这是因为陆地植物所含的蛋白质比海洋浮游植物低得多。由浮游植物到植食性动物,生态效率约20%左右;由浮游动物到浮游动物捕食者,约15%;由低级肉食者到高级肉食者,约10%。
生物的进化,从单细胞到高等动物以至人类,都是沿着改善获能效率和增加获取能量的途径进行的。在生物进化的过程中,生产者和消费者各自的进化水平是相匹配的。化石的分析结果证明,甲藻的出现时期与鱼类的最盛时期相吻合,而硅藻的出现时间大体与鲸类的出现时间相同。J.H.赖瑟(1969)指出以硅藻为基础的食物链要短些,而以鞭毛藻为基础的食物链要长得多。鲸处于以硅藻为基础的食物链上,因此能更好地取得能量,弥补其个体大因而能量消耗也大的不足。
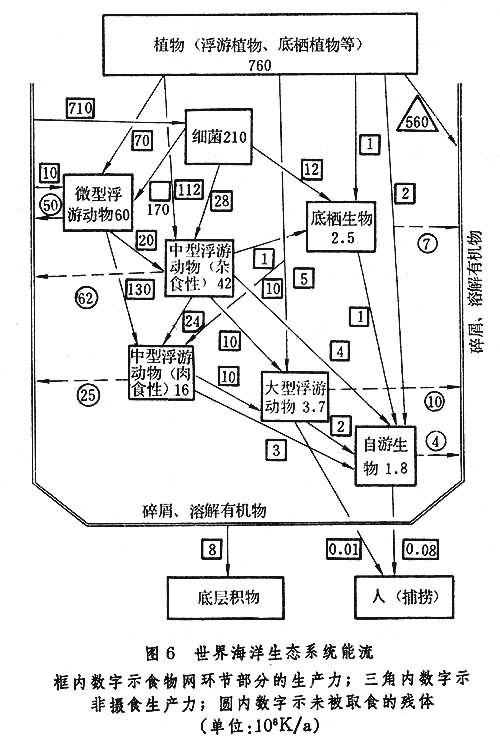
虽然整个海洋是一个连续的整体,不同海域之间没有明确的界线,但根据环境条件的差异,根据生物和环境的特点,可将海洋生态系统分为红树林、河口、浅海、潮间带、上升流、大洋区生态系统,或浮游生物、底栖生物、游泳生物生态系统,或某某海区生态系统。目前有关生态系统的知识还很贫乏,研究工作比较集中于上升流、河口、红树林和几个海区的生态系统。概括已有的资料,已建立几个简单的生态系统模型(图6)。建立模型的目的,是为了更好地表达生态系统内部各种因素、成分之间的复杂关系,帮助人们预测在生态系统遭到各种压力下可能发生的变化,从而达到管理生态系统的目的。
深海生物群落
深海由于压力大、食物少、没有光线和温度低,因此在生物的种类组成、分布格式、个体结构和代谢等方面均有其特点。
深海生物能忍高压。虽然有些浅海生物也能忍受较高的压力,比如附着在潜水器表面的生物,如绿管浒苔、石莼、总合草苔虫、紫贻贝、布纹藤壶等,在潜水器下降到2000~3000米水深后仍然能存活。但根据生理学试验,600个大气压对大多数浅海生物有致死作用。因此,从垂直分布来看,6000米深度似乎是个重要的分界线。曾有报告,中太平洋的深海沟中的125种动物,有77种是在6000米以上水层所没有见到的。
与浅海生物比较,深海生物一般个体数量少,但种类数相对较多、多样性高。对此有不同的解释。H.L.桑德斯(1968、1973)认为,多样性高是由于食物等竞争造成的。但有的学者却认为捕食是关键。较多的调查结果表明,深海生物的多样性仅仅发现在2000~3000米水深处,而5000~6000米以下的海底,生物的多样性并不高。
为适应食物少和黑暗的环境,许多深海鱼类,口部相对扩大,骨骼肌肉减少,且有发达的发光器官和结构。
深海生物一般代谢作用和生长都很慢。据估计,深海的贝类,长到8毫米大约需100年的时间。曾有一只潜水器掉进深海中,经10个月后从1540米处打捞出来,发现放在桌上的三明治仍然完整无损,这表明细菌的作用非常缓慢,但也有例外。
1977年,美国伍兹霍尔海洋研究所曾用深海潜水器“阿尔文”号在加拉帕戈斯群岛以东300公里,水深2500米处进行调查,调查区域是海洋板块形成区,发现从地下喷出泉水,泉水口附近水温高达20℃(没有热泉处的海水是2℃)。在喷出孔附近有丰富的生物群落。其中有个体30~40厘米的贝类,这种贝一年可长4厘米(约比其他深海底的贝类生长速度高500倍),将壳打开,可见到内有血红蛋白(一般软体动物是血蓝蛋白)。还有一种具长栖管的须腕动物,管的直径10厘米长可达30米。此外,还有许多腔肠动物、环节动物、甲壳动物。
深海底栖生物的食源可能包括由上层水中下沉的碎屑和溶解的有机物质,以及化能合成细菌通过氧化硫化氢取得能量而制造的有机物。目前认为,后者是最主要的来源。因为从地下喷出的热水含有大量的硫化氢(30~40ppm),硫磺细菌利用氧化硫化氢所获得的能量将水中CO2合成碳水化合物。海底硫磺细菌实际上起着与浅海水域光合植物相同的作用。即硫磺细菌是深海海底的生产者。这说明,生产者的能源不仅可来自太阳,而且还可来自地球的内部。这是一个重大的发现。根据这一论点,须腕动物的营养问题可得到解释:这种动物没有口,也没有消化道,但在体内有大型的腔,称为营养体,细菌在腔内大量繁殖,动物的触手可吸收无机物供细菌之需,而细菌则合成有机物供动物之用。动物和细菌营互利共生关系。