[拼音]:zhiwu kuangzhi yingyang
[外文]:plant mineral nutrition
植物对矿质养分吸收利用的过程。矿质元素广泛参与植物代谢的各个环节,其中很多元素又是植物的结构物质或植物体内一些重要化合物的组成成分。当这些矿质元素缺乏或比例失调时,植物就会因代谢受到干扰、破坏而呈现相应的病征。植物矿质营养的研究可为栽培作物的合理施肥提供理论基础。
必需矿质元素
植物的必需矿质元素有多种,按需要量的多少可分为大量元素(氮、磷、钾、钙、镁、硫)和微量元素(铁、硼、锰、锌、铜、钼、氯等)两大类。此外,还有一些无素,如硅可增加水稻茎叶的硬度,使叶片更直立,提高光能利用率,增强组织对病虫害的抵抗力;钠对于某些盐土植物和四碳植物有重要作用;钴可促进豆科植物固氮等,证明它们只是某些植物而不是一般植物所必需的,可称为有益元素。大量元素或微量元素都具有重要的生理功能:
(1)植物细胞的组成成分。如氮为蛋白质的组成成分,后者又是原生质的基本成分。硫是蛋白质中胱氨酸、半胱氨酸及蛋氨酸的组成成分。氮和磷又都是作为遗传信息的载体核酸的组成成分。硫脂、磷脂和蛋白质共同形成了生物膜的基本结构,所以氮、磷、硫都是细胞器及新细胞形成所必需的。最重要的光合色素叶绿素也是含氮化合物。
(2)参与能量代谢。如磷是高能化合物腺苷三磷酸(ATP) 的组成成分。在光合作用的光合磷酸化过程和呼吸作用的氧化磷酸化过程中,从腺苷二磷酸(ADP)与无机磷产生ATP,并将能量保存在ATP的高能磷酸键中,当ATP水解时,其所释放的能量即可供各种生命活动之用。
(3)参与酶、辅酶和一些电子递体的组成。所有的酶都是蛋白质,因此都含有氮。此外还有一些其他元素是酶的组成成分,如铁是细胞色素氧化酶、过氧化物酶和过氧化氢酶的组成成分,这些酶参与呼吸代谢。光合链中的电子递体铁氧还蛋白(Fd)、细胞色素 f和细胞色素b6亦为含铁化合物。催化硝酸盐还原的硝酸还原酶则含有钼。有的元素是一些酶作用的活化剂,如 Mg2+是光合作用中核酮糖二磷酸羧化酶及C4途经中丙酮酸磷酸双激酶的活化剂,又是细胞内转移磷酸基的酶的活化剂;K+是丙酮酸激酶、琥珀酰辅酶A合成酶、果糖-6-磷酸激酶和淀粉合成酶等的活化剂。因此上述元素均为正常代谢所不可缺少。由于缺钾时植物组织内丙酮酸激酶的活性即降低,在外观上尚未出现可见缺钾症状时丙酮酸激酶的活性即受影响,这种酶的活性可作为钾营养状况的一种指标。
(4)调节功能。植物细胞液中的磷酸盐、钾盐和镁盐等有调节渗透平衡的功能。磷酸盐还有缓冲作用,是使细胞液能保持一定pH的因素之一。K+在平衡细胞内有机酸阴离子如苹果酸阴离子及无机阴离子如 SO厈及C1-的负电荷中有重要作用。K+和 C1-可调节气孔开闭。Ca2+与调钙蛋白相结合可调节一些酶的活性。如烟酰胺腺嘌呤二核苷酸(NAD)激酶只有在Ca2+、调钙蛋白及核苷酸底物存在时才呈活化态。钙还涉及植物细胞衰老的调节。在衰老过程中,膜的透性增加而区域化降低。钙则可维持膜的完整性,调节膜的透性,从而阻止衰老。硼可促进光合作用中形成的糖从叶向花、果实、茎与根等器官输送。花粉萌发和花粉管伸长也需要硼,缺乏时影响种子及果实的形成。锌为生长素的前体──色氨酸形成所必需,缺乏时会影响生长素的形成,果树小叶病就是缺锌的典型病症。
离子的吸收
植物吸收离子的主要器官是根;此外,幼嫩的茎、叶也有一定的吸收功能。根从土壤中吸收离子需先与土壤中的离子接近,这可通过几种方式:
(1)扩散。在根际附近土壤溶液中的离子沿浓度梯度扩散到根的表面。
(2)根系伸展。根系在土壤中伸展,特别是根毛区,扩大了根的吸收面,同时由于根系生长对根际附近土壤产生挤压作用,使其容重增加,矿质离子更为集中,这些都为根系截获周围的离子创造了有利的条件。
(3)离子交换。根呼吸释放的CO2溶于其周围土壤溶液中,形成H+和HCO婣。H+扩散到土壤胶体表面,即可与土壤胶体所吸附的阳离子如K+进行交换,使H+为土壤胶体所吸附,而将K+交换到土壤溶液中。土壤溶液中的离子可与根表面吸附的离子相交换,又可与土壤胶体吸附的离子相交换,通过这种离子交换的方式,根即可吸收土壤中的离子。此外,当根和土粒接触、根表面所吸附离子与土粒表面吸附的离子的振动空间相重叠时,二者所吸附的离子也可直接交换,这种方式称为接触交换。在根尖到初生根一段的表面有一层粘凝胶,为根分泌的糖蛋白,其中含有游离的-COO-及其他亲水性物质。这一粘胶层可增加根对土粒的接触面,并可吸附较多的离子,促进根对离子的吸收。离子从根表面进入根内则可通过被动吸收,亦可通过主动吸收。
被动吸收吸收过程中不消耗自由能,离子顺电化学势梯度进入细胞。电化学势梯度包括化学势梯度和电势梯度两方面。离子的扩散速度就决定于这两种梯度的大小。在细胞内由于细胞壁中的果胶、蛋白质以及质膜中蛋白质的分子中都有羧基 (-COOH),羧基离解带负电荷(-COO-H+),因而常具有较大的非扩散性负电荷,可从介质中吸附阳离子,使其中的阳离子浓度高于介质中的浓度。细胞内阳离子浓度梯度的大小取决于化学势梯度与电势梯度的相对数值大小,这种膜内外离子浓度不相等的平衡状态称为唐南平衡(图 1)。它是一种特殊的积累离子的现象。
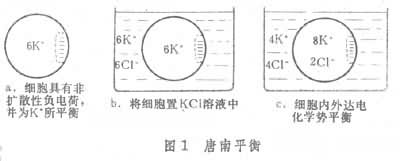
在根外溶液与根内质外体之间溶质可自由扩散,并达到动态平衡。质外体是指原生质以外的结构或空间,又称自由空间。在幼根内,质外体包括表皮和皮层内的细胞壁及细胞间隙这些质膜以外的细胞结构和空间。由于内皮层细胞壁有木栓化的凯氏带,水分和溶质都不能通过,故阻碍溶质的扩散,但在内皮层具有通道细胞的情况下,或在根尖新形成的内皮层细胞尚未木栓化时,质外体即可越过内皮层。此外,通过质流(集流),根亦可吸收土壤溶液中的离子。质流现象是指植物蒸腾作用造成从叶片沿茎、根到土壤的水势梯度,使水由土壤经根表面进入根内,水中的溶质亦随水流进入根内。
主动吸收吸收过程需要消耗自由能,溶质逆电化学势梯度进入细胞。载体学说和离子泵学说是当前对主动吸收的主要学说。
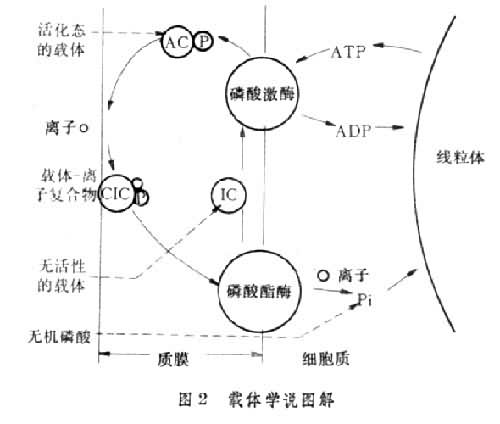
载体学说(图 2)认为生物膜内有能选择性地载运离子通过膜的分子,称为载体,它能在膜内扩散。从无活性的载体转变为活化态需要 ATP。当活化态的载体转移到膜的外表面时,就可与其有亲和力的离子形成载体-离子复合物,然后此复合物扩散到膜的内表面,通过磷酸酯酶的作用使其释放出磷酸基,因而载体失去其对离子的亲和力,成为无活性的状态,同时离子即释放到相邻的介质如细胞质中。无活性的载体通过磷酸激酶的作用又变为活化态,又可在膜的外表面与离子形成复合物。载体可反复利用,因而少量载体即可载运大量离子。在一定浓度范围内,根对离子的吸收随外界介质中离子浓度增高而加速。这个现象可以解释为载体结合离子的部位有限,达饱和后离子吸收速度不能再提高。根对离子吸收的选择性,也可用不同的离子分别有不同的载体结合部位来解释。形成活化态载体需要消耗的能量来自膜上的 ATP。根的质膜和液泡膜中都存在大量分解ATP释放能量的ATP酶。
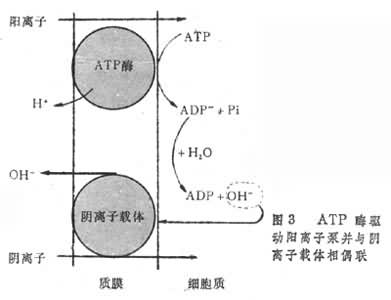
离子泵学说(图3)认为膜内ATP水解,导致阴离子ADP-和阳离子H+的产生。H+释放到膜外,ADP-留在膜内,因而产生跨膜的质子梯度和电位差。细胞内H+浓度较低,电位较负,就使阳离子与H+交换而被“泵”入到细胞内。另一方面,ADP-与H2O反应,增加了细胞质中OH-的浓度,OH-驱动阴离子载体,引起阴离子的吸收。阴离子主动吸收亦需消耗能量,是否直接涉及到ATP酶的作用,还不十分清楚。
根吸收离子可以是主动的,也可以是被动的。两者可以同时进行,但主动吸收常占主导地位。
离子在植物体内的利用
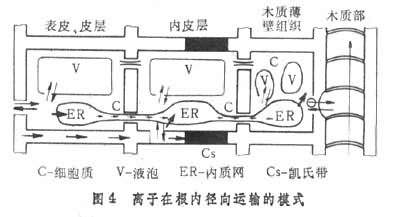
根从土壤中吸收的离子可通过质外体和共质体途径,从根表皮运输到根木质部的导管(图 4)。共质体是指植物体内的原生质通过胞间连丝相互连接而成的整体;进入导管后即随根压流和蒸腾流从根运输到地上部。在运输途径中,木质部输导组织周围的细胞可主动地从导管溶液中选择性地吸取养分,然后运到相邻的其他细胞。当组织衰老时,所含养分可部分地向正在发育的幼嫩部位转移,如老叶中的含氮化合物经降解后运往正在发育的果实及种子中,或运到幼叶及幼根中,形成新的含氮化合物而被利用。磷、钾、镁也很容易再被利用,通过韧皮部的筛管运输。根外追肥时喷在叶面上的养分也通过筛管从叶运到植株的其他各部分。