[拼音]:wuronghe shengzhi
[外文]:apomixis
一种代替有性生殖的不发生核的融合的生殖。在同一个种中,往往有性生殖和无融合生殖可以同时存在。同一种植物可以在某一地区进行有性生殖,而在世界其它地区进行无融合生殖。无融合生殖在植物界是普遍存在的,在藻类和蕨类植物中也有无融合生殖,但在苔藓和裸子植物中迄今却很少报道。本条目只就被子植物的无融合生殖情况做一个简要的叙述。无融合生殖在被子植物中有下列4大类:
营养繁殖
见植物营养繁殖。
在减数的胚囊中的无融合生殖
包括 3种形式:单倍体孤雌生殖,单倍体单雄生殖和无配子生殖。单倍体孤雌生殖和孤雄生殖在正常的单倍体胚囊中,由卵细胞或由雄配子不经融合独自分裂产生胚,它们分别称为单倍体孤雌生殖和单倍体孤雄生殖。这两种情况在自然界是罕见的,曾经记载的例子多数是以远缘花粉授粉诱导产生或是由于冷处理或热处理的结果。单倍体孤雌生殖和孤雄生殖不能产生后代,但单倍体通过人工染色体加倍,可得能育的纯合二倍体植株,因而在育种实践上有很大的意义(见植物实验胚胎学)。
在未减数的胚囊中的无融合生殖
包括两种形式:
二倍体孢子生殖又称种细胞无孢子生殖,由二倍体的胚囊中的卵细胞不经受精发育成胚。卵细胞一般自发地发育(不需要传粉的刺激),而且这些植物的花粉在发育过程中减数分裂不正常。不过,也有一些种须要假受精,即须要传粉的刺激。例如高山早熟禾。这种无融合生殖方式的胚囊的发生有下列 3种不同情况:
(1)大孢子母细胞分裂早期维持不配对的染色体,后来形成再组核,经有丝分裂发育为二倍体的胚囊。例如齿叶苦荬菜,一年蓬和金光菊(图1)。
(2)大孢子母细胞分裂早期染色体不配对,而后形成一个再组核,经正常的有丝分裂形成二分体,由合点端的一个二分体细胞产生胚囊。例如白花蒲公英(图2)。
(3)大孢子母细胞休眠期较长,而后进行有丝分裂,直接产生二倍体胚囊。例如腺泽兰以及拂子矛属、蝶须菊属、早熟禾属中的若干属(图3)。因此属二倍体孤雌生殖。
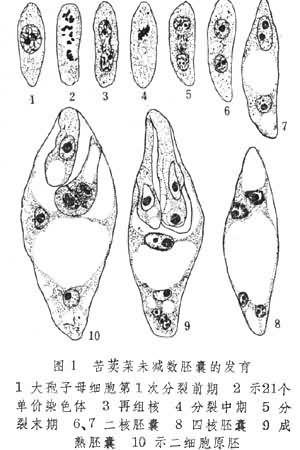
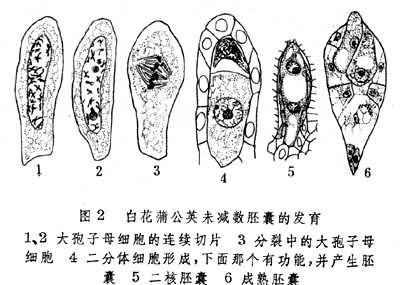
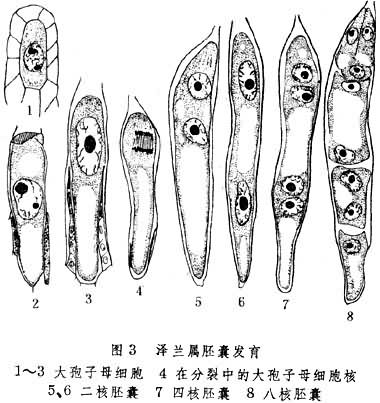 体细胞无孢子生殖
体细胞无孢子生殖
胚囊是由珠心细胞起源,在这种二倍体胚囊中的卵细胞以孤雌生殖的方式(二倍体孤雌生殖)发育成胚。偶尔有由胚囊中其他细胞产生胚的。
在体细胞无孢子生殖的种中,大孢子母细胞早期退化或者经减数分裂形成一个正常的胚囊,但迟早被无孢子生殖的胚囊取代。这种无融合生殖的种需要假受精,花粉的发育是正常的。体细胞无孢子生殖见于极地早熟禾及还阳参属、银胶菊属、委陵菜属、毛莨属、悬钩子属等。
从珠心或珠被细胞产生胚
也称不定胚发生。不定胚是由珠心或珠被细胞直接发育。在发生不定胚的胚珠中,胚囊是正常发育的,预定形成胚的胚珠细胞进行活跃的分裂形成细胞群,并逐渐推进至胚囊中继续发育为与合子胚相似的胚状体。
具不定胚的种可以有两种情况,一类形成胚乳不需要受精。例如Alchornea ilicifolia,Euphorbia dulcis和Opuntia aurantiaca。在这些种中减数的卵细胞早期退化,而不定胚从接近胚囊的珠心细胞形成。胚乳自发地发育。另一类如柑橘属的许多种,胚囊中的卵细胞正常受精发育成胚,同时由珠心细胞直接形成不定胚。因此胚囊中间同时存在合子胚和不定胚。一个胚珠中产生的不定胚常常不只一个,甚至形成多胚(见多胚现象)。柑橘类的种子中有时可达30个以上大小不等的胚。