[拼音]:qianxi
[外文]:migration
动物周期性地往返于不同地区之间的远距离移居行为,例如候鸟的迁飞和鲸的洄游。鸟兽离巢觅食早出晚归,不论外出多远,这行为都不属迁徙范畴。这里远距离一词是相对于动物的身体大小和行动能力而言,可以由数十里直到近万里,但它们在其间往返的地区在气候、食物、天敌等生态因子上必然有所不同。周期性一词也只是指整个种群而言,王蝶至秋季可飞行二、三千公里到南方越冬,成虫寿命不长而长途飞翔需时较久,故最后返回原地者已属子代,就个体而言可能并未经历一个完整周期。此外还有人用迁徙一词来概括动物的一切长途移动,但本文采用较严格定义,只限于返回原地的移居行为,不讨论巢区扩张以及一切一次性迁出的行为。另外还有一种漂泊的生活方式,例如澳大利亚干旱区的野鸭巡回觅食,当地雨水稀少,且分布不规则,下雨地区一时食物较为丰盛,野鸭即游荡其间。野鸭栖息的地区变换不定,这种行为也不属于迁徙的范畴。地球上除赤道外均有明显的季节变化,食物、天敌等等亦随之增减波动。而高纬度地区的严冬低温非一般生物所能适应;有些植物以地下部分或种子越冬,有的动物栖息洞穴之内减少外出或甚至冬眠,有的动物则向温暖地区迁移,特别是在具备迅速行动能力的鸟类中就发展出长途迁徙的行为。
迁徙的类型
迁徙行为主要见于鱼类、鸟类和哺乳类,一般以一年为周期,其水平距离常跨越不同的温度带,且除幼体外主要采取主动移动方式。但有些低等生物,如海洋浮游动物有一种昼夜升降的垂直移动现象,习惯上也称之为迁徙,但这些行为不够典型或根本不符合前述定义。这些动物夜间升至水面表层取食,白昼则下降,这有助于避敌。光强和水温的变化可能是移动的触发因子。但其性质类似鸟兽觅食,并非两地往返移居。再如蝗虫,当虫口拥挤食物短缺时会出现一类群居型个体,它们群集起飞,主要被气流裹挟至低气压易降雨的地区。遇适当的潮湿环境条件蝗虫即停止移动,而且蝗虫发育成熟后即交配、产卵以至死亡。这是一种播散现象,也不是真正的迁徙。
迁飞空域迁徙的别称。空域辽阔易于定向,除异常天气外极少障碍,飞行速度也远比在水、陆迁徙为快,故在此出现最发达的迁徙行为如鸟类的迁飞。飞行最远的当属北极燕鸥,它们繁殖于欧洲、亚洲、北美洲的极地部分,但秋季却飞越重洋至南极浮冰上越冬,航程近18000公里(见图)。
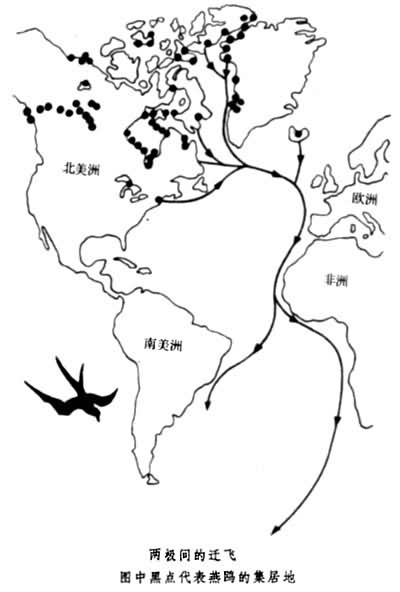
跨越赤道前平日赖以定向的太阳位于自己的南方,越过赤道后太阳却移至自己的北方,且旅途中日照长短及气候条件变化极大,但它们仍能准确地辨明时间和方向。一般鸟类的迁飞是往返于繁殖地区和越冬地区之间,两地都要求在栖居期间食源充足、天敌少。局部的气候变化和食源消长往往是迁飞的动因。因此,同一物种中可能同时存在迁飞和不迁飞两种类型,视栖息地区的情况而定。例如,西欧的环境条件稳定,当地的紫翅椋鸟不迁飞,而东欧越冬条件较差,那儿的椋鸟迁飞至西欧或北非越冬。在昆虫中也有真正的迁飞,例如,西方五月鳃角金龟成长后即迁至林区取食,待性成熟后再返回原地生殖,往返距离不大,但行为规律,且一生中可重复数次。再一类型昆虫迁徙是由繁殖地区至冬眠或夏眠地区,越冬或越夏后再返回原地排卵。王蝶可列入此型,它们在夏季生活于加拿大和美国北部,秋季南下至美国南缘冬眠,迁飞距离达3200公里上下。次春返回时速度较慢,出蛰后及途中可数次繁殖,故返回者主要为子代。此外,飞行哺乳动物蝙蝠也有类似的迁飞行为,如北美红蝠夏季栖息于加拿大和美国交界一带,10~11月间南飞至美国南部准备冬眠,甚至有跨海飞至百慕大群岛者。
洄游即水域迁徙。水体中较难定向,辽阔海域虽少障碍物,但行动难易全看行动方向与水流方向是否一致。大多数鱼类都有洄游行为。洄游大致有3类:
(1)生殖洄游。指鱼在性成熟后由取食或越冬地区游向产卵场所的移动;
(2)索饵洄游。生殖洄游之后,为了弥补身体亏耗需要大量捕食而进行的洄游。对雌鱼意义更大。因饵料生物数量的变动以及饵料生物本身也可能洄游,所以鱼类必须洄游以追逐食源。路程可长达几千里;
(3)越冬洄游。喜温鱼类每当冬季降温时即游向温暖地区越冬,因洄游期间少食或不食,故常在索饵肥育之后进行。生殖洄游还有往返淡水海水之间者。平日栖居海内,至生殖季节溯河返回淡水内产卵者称溯河鱼类,如多种鲑鱼,包括中国的大麻哈鱼。幼鱼阶段时它们生活于淡水,两三年后入海。成鱼在海中度过两三冬再溯河而上,一路上顶急流、越浅滩、逆河千里,故常在产卵后死亡。标记实验证明,大西洋鲑鱼可返回原生长所在的河流。进一步实验证明,它们主要依靠嗅觉来辨认源流。但难以理解的是,幼鱼降河时的嗅觉记忆竟能维持数年之久。还有一类鱼称为降河鱼类,它们大部时间栖居淡水,只是在生殖季节入海产卵。如鳗鲡生于大西洋马尾藻海水深400~700米处。幼体被墨西哥湾流带至北美洲东岸或欧洲西岸。两三岁时发生变态,然后于春季溯河返家。在淡水中长至10~15龄再降河入海,回到马尾藻海产卵后死亡。除鱼类外,能长途洄游者还见于海兽,如南极鲸类常至非洲西北岸的大西洋海域或印度洋的亚丁湾和孟加拉湾等处越“冬”(相当北半球夏季),而部分北极蓝鲸也可抵达印度洋越冬。两栖类和爬行类中少有能远途洄游者,但海龟却属例外,绿龟生于大西洋西部的阿森松岛,却远渡重洋至巴西东岸海域取食,两地相距约2000公里。
陆地迁徙
陆地障碍较多,动物难以直线行进,而且陆行速度不快,消耗却较大,故陆地迁徙较不发达。显见的例子,如北美驯鹿夏季取食于北极圈内冻原上,7月开始南下然后在泰加林中越冬,至次春2~3月间再度北移,每年都走同一路线。非洲的斑马在干湿季节变动时追逐水草,可游动千里以上。此外还有一类垂直迁徙:某些小动物在山区高处繁殖,但当天气转寒时下至山脚或平原越冬。它们移动的路程虽短,但随季节往返于两种生态环境之间,这种行为仍属典型的迁徙。
迁徙的机理
目前对鸟类迁飞研究得较为深入。迁飞是一些鸟类(候鸟)生活史中的一个组成部分,个体发育中各个阶段按照一定的内在程序陆续展现,同样,迁飞也可能决定于某种内在节律。这内在节律又必须同外界的自然节律相吻合,这样才有利于鸟类在环境中的生存和繁殖。远在迁飞之前,很多鸟类就在体内积蓄脂肪,体重可能比繁殖季节增加一倍。这种代谢改变受甲状腺控制,而甲状腺的发育又受垂体的影响。实验证明,日照时间的延长可以通过垂体促进性腺及甲状腺的发育,改变鸟类的代谢状态。
日照时间的变化不仅指明起飞的时刻,还可影响航行的取向。曾有人以小靛蓝彩鹀作实验,将它们分为两组,接受不同的人工光照。一组接受日渐减短的光照,这相当于秋季(南向)迁飞前的情况。另一组接受日渐加长的光照,相当于春季(北向)迁飞前的情况。然后将两组同时置于天文馆内,在屋顶投照春季星空景象,结果第一组南飞,第二组北飞。不过日照时间代表的是日历时间,却不能指明外界的生态条件,如果仅仅按日历时间起飞而不顾当时气候如何以及食物是否充足,则有可能导致伤亡。故很多鸟类还依靠其他环境信息,例如气温、气压以及食源情况等等。
鸟类常在迁飞前表现烦躁不安,经常朝向未来迁飞的方向,这个现象称为迁飞前躁动。大部分鸟类在迁飞时有集聚倾向,聚成或大或小的群。一般说,食肉鸟类如猛禽和食昆虫的雀类在结巢及猎食期间高度分散,但在迁飞前也趋向集群。结队迁徙可能有助于更准确地定向和减少群体因克服阻力而消耗的能量。
鸟类迁飞的本领惊人,北极燕鸥一年一度往返两极之间,单向行程近2万公里。实验表明,许多鸟类每年可准确地返回到同一地点。金斑鸻自阿留申群岛经海面直飞夏威夷群岛并无地形标记可循,有关机理见定向和定时。但一般鸟类多沿河流、山脉、海岸线飞翔。欧洲白鹳自欧洲至非洲的迁飞有两条路线:西线越直布罗陀海峡,东线则经土耳其、叙利亚、以色列和西奈半岛等地,两条路线都似乎在避免跨越地中海海域。鸟类迁飞时航速大于平时的时速,常在70公里上下。飞行高度因物种而异,高者可在4000米以上,但高山峻岭如阿尔卑斯山成为多种鸟类的迁飞障碍。
鸟类有昼飞者如鹈鹕、鹳、猛禽和燕子等;有夜飞者如水禽、杜鹃、鸫等,并主要在前半夜飞行。一般鸟类多在路间停歇,寻食和休息可占去大部分时间。还有的野鸭飞至安全地带换羽(称为换羽迁飞),然后再飞往越冬地。
遗传与进化
椋鸟的幼鸟至迁飞季节就表现出上述的生理反应,包括迁飞前躁动,这说明迁飞在很大程度上是由遗传决定的。因此问题变为:迁飞行为是如何在生物进化史中出现的。目前所见到的迁飞行为都不外寻求一个温度适宜、食源丰富、天敌稀少的场所。不论是取食(或称索饵),还是生殖,还是越冬,都离不开这几个要求,只是在生殖季节,这方面的要求更加强烈。纬度越高,季节性的气候变化越明显,食源也随之剧烈波动。在这种情况下,具备高速航行本领的动物中就发展出远距迁徙的现象。也可以说,地区资源的季节性变动导致了不同气候区的承载力的消长,再结合动物在生殖和越冬时期的对环境需要的不同,就造成动物的季节性迁徙。因此,迁徙主要见于资源波动较大的地区。例如北方寒冷地区冬夏温差太大,而热带稀树草原干温季节也气候悬殊,某些水鸟即迁徙其间。对比之下,热带雨林虽食源丰富,但因供应稳定,生态位保持基本饱和,故鸟类基本不迁徙到热带雨林。有关迁徙的具体进化过程目前只有一些推测。可能一开始只是一种巢区扩张,有的动物可能在寻食时探索得更远些,还可能有些动物偶然被气流或水流裹挟到远方。原地的资源短缺或捕食者追猎都可能迫使动物放弃旧居外迁。最初移动距离较近,随后越移越远。第四纪冰川可能是促使很多鸟类远距迁徙的重要原因。例如,鸻、野鸭的繁殖地靠北,可能气候转冷后它们被迫迁至南方越冬。但黄鹂及雨燕原栖居热带,后来随着冰川后退,大陆转暖,它们在春季也转至较北地区繁殖。不过,对鳗鲡那样漫长迂回的迁徙,却没有得出满意的解答,或怀疑这与大陆漂移有关,但尚未得证实。
参考文章
- 郭庆兰:爱,绝不迁徙保健养生
- 中国历史时期人口的分布和迁徙(distribution and migration of population inChinese history)中国地理
- 为什么日耳曼人要大迁徙中国史
- 什么是贷款迁徙分析或信用风险迁徙分析?银行业务
- 鸻形目的中国历史记载、外形特征、外形特征、环志与迁徙、分类地位、地理分布、化石、保护现状、亚目、鸻形目大全鸟类知识
- 短尾鹱的分类地位、基本资料、外形特征、生活习性、生长繁殖、动物迁徙、最新发现鸟类知识
- 白头鹞的基本信息、地理分布、外形特征、叫声、生活习性、求偶、生长繁殖、迁徙、保护现状、地理分布、食物、亚种鸟类知识
- 黑眉拟啄木鸟的物种信息、分类地位、外形特征、叫声、地理分布、品种、生活习性、食性、迁徙、保护现状鸟类知识
- 红喉潜鸟的基本资料、外形特征、地理分布、生长繁殖、动物迁徙、保护现状鸟类知识
- 海南蓝仙鹟的简介、外形特征、地理分布、迁徙、生境、食性、生活习性、生长繁殖、野外鉴别、生存状况、相关条目鸟类知识