[拼音]:dingxiang
[外文]:orientation in space
动物主动调整其身体或身体某部分的空间位置的行为。如青蛙伸舌粘取飞虫的取食行为,以及候鸟往返数千里飞翔于越冬及繁殖地的迁徙,就其需要精确地确定方向而言,都属于典型的定向行为。定向行为的对象可为外物,如鸟兽类使所筑巢穴有一定朝向(如避风朝阳)。空间位置可以指静止的位置,如动物休息时哪一部分贴附于基底都有定规。又如在阳光下,蜥蜴常根据外界温度而采取一定的体位方向使所吸收的日光足以保持体温大致稳定。再如猛兽隐藏树丛中准备伏击过路动物,必须维持朝向一定方向的警觉待动的姿态。但动物还要不断地活动,要觅食、求偶、避敌等等,这都存在定向问题。动物的捕食动作不仅要敏捷有力,而且一定要对准目标。飞鸟四出寻食路途迂回,甚至会遇到风雨,但却能顺利返巢。这一切的关键在于动物能辨别方向。在定向行为的全过程中随时都要辨别方向,校正可能发生的偏差。
方向与定向
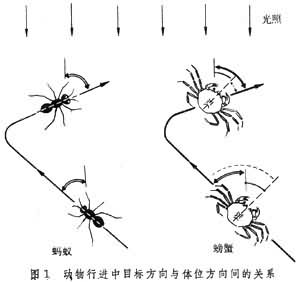
动物的静止姿态和活动路线大都有明确取向。动物的四出觅食似无目的,但实际上有内在规律,或循气味、声响等线索,或采取某种程序按区搜索。例如蚂蚁在植叶上常是沿某一边线(或左或右)循序查遍各区。再如海边的招潮蟹的巡视路线很象一朵花,自同一点出发沿辐射方向外出,画一个小弧回来,再沿另一辐射向外出,如此循序绕遍几个方向。
方向是就一定的参考系统而言。可能是针对一个点,例如捕食的目标、求偶的对象、返归的巢穴等;也有的是就某个方面而言,例如沿重力线有上下之分,上下本来只是相对而言,但若划定一个界面(如地面或水面),便有了绝对的分界。不过在一般描述中总采用一定的坐标系统。在左右对称的动物中,可以利用3组剖面将身体分为头尾、腹背和左右几个方面。在横剖面上,以头尾轴作中心还可分内外方向。至于动物在环境中的位置,则一般总是采取地理坐标,即东西南北方位。
体位的定向动物不活动时的体位大多是相对于重力线、基底或外界光线而保持一定方向。在倾斜的地面上,体重较大的动物总力图保持其垂直轴与重力线一致。但重力对微小动物的影响就不如基底的影响大。例如家蝇在顶板上和地面上的行动同样自如。许多水生动物因水浮力的关系,受重力的影响较小,而顶侧来的天光成为其重要的定向参考因素,这些动物总是保持背向天光的体位。再因为它们背侧皮色较深而腹侧较浅,所以与顶侧天光配合,还有利于隐形避敌。
大多数动物行动时,身体头尾轴与路线方向一致;但螃蟹行动时,不管路线如何迂回,其身体却保持与环境成一固定角度,因此身体方向与路线方向并不一致(图1)。
 行动路线的定向
行动路线的定向
对于短距离行动,地形或气味标记,便可引导动物直达目的地;而远距离航行则需要其他线索导向。从理论上讲,地球上两点相距不论多远,要在其间直线航行只需要知道方向和距离。只要取向正确,动物航行中所消耗的体力可有助于估计走过的距离,而局部地形则有助于辨识是否已到达目的地。因此定向就成为关键问题。仅仅知道东西南北各在哪方不行,要知道的是目的地在自己的哪一方。
对于人类飞行来讲,只需要知道起飞地和目的地各自的经度纬度,便可在地图上找出方向和距离。但动物无相应机制。目前所能肯定的只是很多动物能以太阳或其他天体为定向的指针,不过这也只是指出动物四周的地理方位,并不能据此判定目的地在何方。对于地表观察者来说,天体位置是随时间而有规律地变动着,因此,还要借助定时能力来判断方位。
定向的机理
动物的行为都是内外因子的综合结果,内部生理状态产生行为的动因,外在环境因子则触发具体行动。对于定向行为来说,还存在导向因子。例如小鼠见猫急奔回穴,猫是奔跑行为的触发因子,洞穴则是导向因子。而猫捕鼠时,鼠对于猫来说,既是触发因子,又是导向因子。
感觉辨向准确地辨向有两个条件,一是要求来自导向因子的信息能沿直线行进。光线在均匀介质中是直线传播的,因此视觉成为高等动物的主要辨向感觉。相比之下,气味随气流变化而改变路线,依嗅觉寻找食源便不很准确。二是要求感官本身具备定向的结构。
动物感官可大致分3种类型:单感受器型、双感受器型和屏幕型,其辨向能力依次加强。例如蜂类寻食时,侧方飘来的香气形成一个浓度梯度,使它的两个触角感受的刺激强度不等,它马上转向刺激较强的一方。但若梯度不明显,每个触角感受的刺激强度没有差别时,它便摇摆身体,探查空间相距较远的两点间的气味浓度差别来定向。前一种情况下,是同时利用双感受器定向;后一种情况是两个触角只作为一个感受器,每一时刻感受到的只是一点浓度,辨向是综合不同时间不同地点所得感觉的结果。但若外来信息是直线传播,某些单感受器也足以辨向。如响尾蛇头部的热辐射探测器深陷在颊部小窝内,小窝口部较窄,仅容一定方向的辐射进入,故用单侧颊窝即可辨向,但双侧颊窝的探测范围在中线方向有些重叠,两者同时使用更有利于瞄准目标。兽类的声感受器也是深陷于耳内,某些兽类的耳廓可灵活移动对准声源,用单侧耳即可辨向,但通常是两耳并用,或因同一相位的声波先到达近侧耳,或因近侧声响较大,从而辨出声源方向。辨向能力最高的器官类型是屏幕型,如眼,一只眼内可有千百乃至亿万个小感受器排列成阵。在节肢动物突起型的复眼上,每个小眼都有方向性,分别接受不同角度的光线;脊椎动物的眼为内凹型,外界光线通过聚光系统一一对应地投照在各个感受器上。复眼另有一个好处,就是可辨别光线的偏振方向。动物眼睛一般成双,当两侧视野重叠时,双眼还有助于探测距离。
来自外界导向因子的刺激,也可大致分为3类:机械的、化学的和辐射的。机械性刺激包括触压觉、平衡觉和听觉等。龙虾的螯肢上覆有一层感觉性纤毛,纤毛的不同排列可以探测不同方向的水流,这些纤毛组成一个屏幕式感觉器官,足以辨别水流方向的种种变化。平衡觉一般是辨别重力方向以及各种加速运动的方向,帮助调整身体姿势。机械刺激中,以波动最有助于定向。仰蝽可根据水波方向发现附近的猎物,这属于低于声频的次声波。高于声频的超声波,方向性更强,借此可在黑暗中辨物。蝙蝠能在深夜或无光线的岩洞中捕捉飞虫,就是靠本身发出超声束再借感官接受反射回的超声波辨向的。其原理近于雷达,被称为声纳。
所谓化学性刺激主要是通过嗅觉感知的各种气味。食物、配偶或天敌都可散发出特殊气味,形成一个自内向外由高到低的浓度场。动物可借浓度梯度辨别气味源的方向。例如,蚂蚁常在取食路途上散布特定的分泌物,供同巢蚂蚁循迹跟踪。另外,气味随气流运动,故逆风而行是很多动物辨向觅食的重要方式。
动物可以感觉远距离发来的辐射,而且定位准确。例如,在人类的视觉范围之外,响尾蛇可察觉微弱的红外辐射;某些昆虫可感受紫外辐射以及光线的偏振方向;有少数鱼类可以在水中产生微弱电场,借此察觉一切游入电场内的其他生物;一些鸟类可以感觉天然磁场,欧歌鸲在天然磁场下采取一定方向,但在附加人工磁场后,即转而采取两者的合力方向。
航行定向一切远出觅食后返巢饲雏的鸟兽都必须具有一定的辨向能力。蜜蜂离巢数百米觅食时,盘绕迂回飞行,而返巢时却可直线飞回,说明它能判断巢在自己的哪一方。也有令人难理解的例子,如鲑、鳗等远距离迁游和候鸟远距离迁飞是如何定向的。人们研究最多的是信鸽,将信鸽运至近千公里之外再释放,不到一天就可返回家。在陌生地点,它开始时只是绕圈飞,但越绕越大,然后便以近于直线的路程返家,直到发现熟悉一些的地形标记时才最后调准路线直抵家中。有些地面动物行走时沿途以分泌物标记,故可以循原路返回。实验证明,细腰蜂可以辨识巢穴周围的地形标记。可能很多动物在日常活动中是依靠辨识地形定向的,如信鸽在接近目的地时是如此。很多候鸟常沿河流、山脉、海岸线迁徙,可能便是把地形作为导航的一个手段。但无论气味标记或地形辨识都只能引导动物重走旧路,而不能解释信鸽在陌生地点被释放时的定向行为。
动物在远航时可能同时采用多种导航手段,在航线的不同阶段重点有所不同。各物种之间采用的手段也不相同,这与它们所具有的感觉能力有关。远航前,动物需要知道的不仅仅是东南西北,而是目的地的确切方向,在航行中还要能保持这个方向,不论是气流、水流或地形障碍使它偏移航向多远,它都必须能及时发觉和纠正。在这方面,地形及视觉辨识只能起辅助作用。有人曾提出,地心引力和因地球自转造成的科里奥利氏力都随纬度而变,动物可能借此判断自身所处纬度。但实验未能证实这个设想。某些动物确能感觉地球磁场,可能以此作为导航手段,但这也只能决定南北方向的位置。
目前比较肯定的是以天体(特别是太阳)作指针的定向航行。实验室内养的椋鸟在春秋迁飞时期即表现燥动不安,当置于四面只有窄窗的笼内,在无云天时经常朝向迁飞方面。但若在笼外用镜子将光亮方向顺移90°,则椋鸟朝向的角度也顺移90°。因太阳始终在运动,以太阳做指针定位,必须知道时间。曾以信鸽做实验,将它囚于密闭室内,室内明暗交替,较外界日夜周期错前6小时。经过一段时间的训练,信鸽体内生物钟提前6小时。然后将实验鸽和作为对照的正常信鸽携至家巢的南方,并于清晨时释放。结果发现对照信鸽朝北飞(返家方向),而实验组却主要朝西飞。这可能因为清晨太阳在东,北方在太阳之左,正常鸽时间感正确,即朝太阳左方飞去。但实验鸽的时间感提前6小时,自觉已至中午,中午太阳在南方,是时北方应在太阳的对方,所以它便向太阳对方飞去,实际是朝向西方。某些鸟类还以星辰为指针。理论上讲,只要找到星空旋转时的不动点,便可知道北极所在,也就定出南北方向。大部分这类实验是在天文馆中做的,利用投照在屋顶的星空影相模拟真实星空。初出巢的小靛蓝彩鹀就会观察星空,注视哪些星辰在星空视运动中相对静止。它系向南迁飞,也即背离北极星。在天文馆中,使屋顶星相不以北极为中心,而以猎户座作中心旋转,发现它选择了背离猎户座的方向朝北方飞去。莺辨识能力更高,实验时只要屋顶上显示出主要星辰,甚至星空并不旋转,它也能定向,这说明莺能辨识星空图像。以上实验都说明,很多候鸟能以天体作指针导航。自然,这决不是唯一的手段。例如很多鸟在密云之下仍可借磁场保持航向。鸟类的平衡器官 (前庭)也可能有助于维持航向,类似于飞机的惯性导航系统。但确定地理方位并不等于确定目标方向,信鸽在陌生地释放后如何找出返家方向仍然没有得到明确答案;在异地航行中发生偏离时又如何纠正也同样是未解决的问题。
遗传与进化
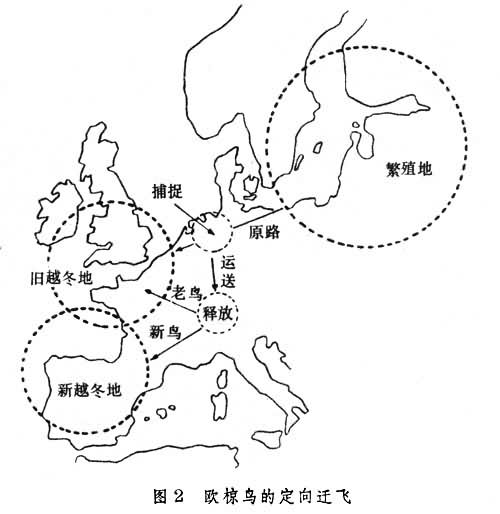
欧洲园莺自幼在笼中饲养,至秋季迁飞时节经常面向西南方(习惯迁飞的方向)。这说明定向行为有其遗传基础。但波罗的海一带繁育的椋鸟的传统越冬地在英格兰和爱尔兰的南部及法国北部。在秋季,当它们途经荷兰时加以截获并套环标记,再送至瑞士释放。这样通过连续几年的实验。结果发现初次飞行的新鸟仍按习惯的地理方向飞行竟飞到西班牙一带去越冬,而飞过一次以上的老鸟却能改变航向,仍飞到传统越冬地(图2)。这表明,学习与经验有助于确定目标方向。
动物的各种行为都有针对的目标,因而都有定向问题。无脊椎动物中,定向能力以昆虫最为发达。脊椎动物中,则以鱼类、鸟类和哺乳类中一些年年远距迁徙的物种表现得最为突出。而且随着环境介质不同,动物演化出不同的定向手段。陆地上,气味标记是较古老的方法。在黑夜,在地穴和海洋中,某些飞行的和水生的哺乳动物演化出超声定向的本领。但定向能力最发达的还要属鸟类。有的候鸟能飞翔万里,具有敏锐的视力足以辨识大面积的地形特征。很多候鸟还具有生物钟,因此能借太阳或星辰指向,甚至在浓云密布的天气中,它还可借磁场来保持航向。