[拼音]:tangzhi
[外文]:glycolipid
含糖的脂质化合物,其糖(链)与脂中的醇基以共价连接。糖脂除参与质膜构成外,还与细胞表面标志及细胞间的粘合等生物学功能密切相关。
糖脂广泛存在于各种生物体中。自然界中的糖脂可按其组分中的醇基种类而分为两大类:甘油糖脂及鞘糖脂。糖基化的甘油醇脂类称为甘油糖脂,或称糖脂酰甘油,存在于动物的神经组织、植物和微生物中,是植物中的主要糖脂,亦是某些细菌,尤其是革兰氏阳性细菌菌膜的常见组成成分。糖基化的鞘氨醇脂类称为鞘糖脂或糖鞘脂,最初发现于神经组织中,以后发现在动、植物的各种细胞膜中普遍存在。其生物学功能远较甘油糖脂复杂。其类型及其与其他各种类脂在组织、细胞中所占的比例因种属及器官而异。尤其是肾中糖脂的种属特异性十分突出。仅含中性糖及氨基糖的鞘糖脂称为中性糖脂。其中只含一个单糖基的称为脑苷脂。半乳糖脑苷脂在脑白质、脊髓及肾中含量丰富,而葡萄糖脑苷脂则存在于非神经组织的细胞膜中。脑苷脂去掉脂肪酰则称为鞘氨醇糖苷。硫酸化的脑苷脂称为硫酸脑苷脂或脑硫脂。含唾液酸的鞘糖脂称为神经节苷脂,因最初在脑组织中发现,误认为是神经节特有的成分而得名,以后证明广泛存在于神经组织以外的各种细胞及体液中。此外,甘油磷脂或鞘氨醇磷脂亦均可发生糖基化。
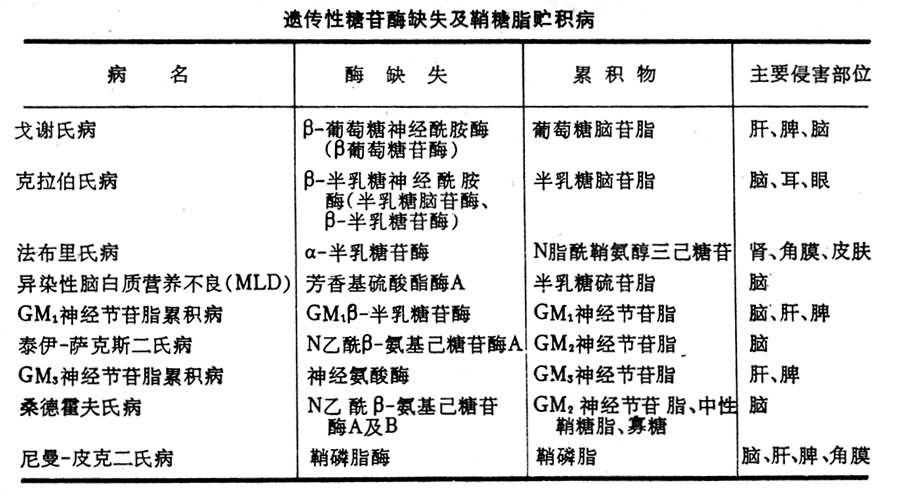
糖脂的生物学作用见表1。
 结构、分类及命名
结构、分类及命名
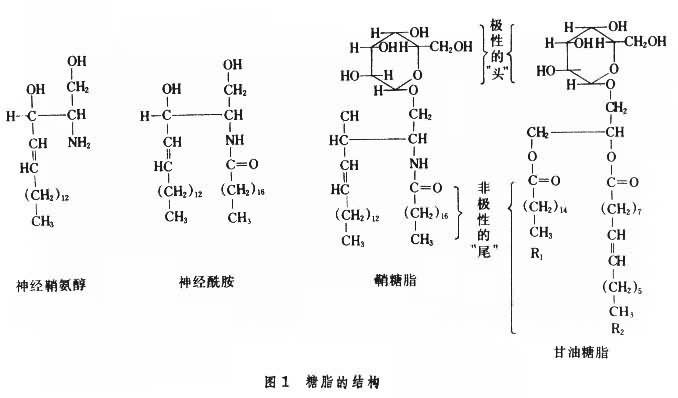
糖脂的组成成分主要是醇、脂肪酸及糖(图1)。有的还有硫酸或磷酸。甘油糖脂的醇为丙三醇(甘油),与脂肪酸形成酯键。鞘糖脂的醇为鞘氨醇(神经鞘氨醇),主要存在于哺乳动物,以其C-2氨基与脂肪酸形成酰胺键,产生N-脂酰鞘氨醇,又称神经酰胺。糖成分一般构成糖链(直链或有分枝)。糖链多以糖苷键与醇的羟基连接,分别形成甘油糖脂或鞘糖脂。有的与磷脂酰甘油或磷脂酰鞘氨醇的磷酸基形成酯键,还有与肌醇磷脂或肌醇鞘磷脂的肌醇形成糖苷键。最简单的糖脂只含一个糖基,如半乳糖脑苷脂及葡萄糖脑苷脂。复杂的鞘糖脂可有20个以上糖基组成的糖链。鞘糖脂中常见的糖基为D-葡萄糖 (Glc)、D-半乳糖 (Gal)、D-乙酰氨基葡萄糖(GlcNAc)及D-乙酰氨基半乳糖(GalNAC),有的还有L-岩藻糖 (Fuc)、D-甘露糖 (Man)及唾液酸(SA)。不含唾液酸的非硫酸化鞘糖脂称为中性鞘糖脂,不带电荷。含唾液酸的鞘糖脂称为神经节苷脂。神经节苷脂在中枢神经系统含量较高,在灰质中的含量相当于白质中的3倍;在非神经组织中含量虽不高,但功能非常重要,是膜脂不可缺少的成分。神经节苷脂中唾液酸的羧基在生理性pH下解离而使整个分子带负电荷。
鞘糖脂的某些糖残基(常为Gal的C-3羟基)可发生硫酸化,成为带负电荷的硫酸鞘糖脂。它们主要存在于神经组织的髓鞘及白质中。例如硫酸化的半乳糖脑苷脂约占脑白质脂类的15%。
神经节苷脂及硫酸鞘糖脂属于酸性鞘糖脂。它们根据鞘糖脂糖链根部的结构可分为7个不同的系,传统命名为GM4、GM3、GM2、GM1、GD1a 、GD1b、GT1a 、GT1b及GT1c。
综上可见,甘油糖脂为糖基化的二脂酰甘油,鞘糖脂为糖基化的单脂酰鞘氨醇。两者各有两条非极性的疏水长“尾”(前者由两个脂肪酰提供,后者由鞘氨醇及脂肪酰各提供一个)。这两条疏水长“尾”可嵌入于膜脂双层,由糖基构成的极性“头”部伸向质膜外,可作为细胞表面标志或抗原决定簇或受体的识别部分。
特性糖脂属于两亲(即亲水又亲脂)性物质。其水溶性差别很大。有的完全不溶于水,如含单糖基的脑苷脂;有的则能以微泡或聚合物形式“溶”于水,如复杂的中性鞘糖脂及神经节苷脂。糖脂分子中由鞘氨醇及脂肪酰烃链构成的疏水性长尾通常嵌入于质膜脂双层的外层,故亲水性糖链仅暴露于细胞外表面,呈不对称分布。
代谢鞘糖脂的转换率在中枢神经系统,尤其在新生儿期,十分迅速。
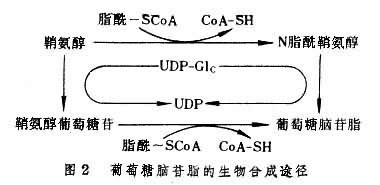
鞘糖脂的合成过程经过多步骤在内质网及戈尔吉氏体中进行。整个合成过程包括鞘氨醇的生成、鞘氨醇脂酰化(神经酰胺的生成)及糖键的合成三个阶段。鞘氨醇系由脂酰辅酶A与丝氨酸缩合并脱羧,再经还原及黄酶催化的脱氢而生成。鞘氨醇可通过与脂酰辅酶 A或游离脂肪酸作用而进行脂酰化生成脂酰鞘氨醇(神经酰胺)。脂酰辅酶A参加的反应由脂酰转移酶催化;游离脂肪酸参加的反应由一种水解酶催化(即神经酰胺分解的逆反应);一般以前者为主(图2)。糖基化的第一个产物为葡萄糖脑苷脂或半乳糖脑苷脂。接受第一个糖基的可为脂酰化的或未脂酰化的鞘氨醇。前者直接生成脑苷脂;后者先生成鞘氨醇糖苷,然后再脂酰化生成脑苷脂。单糖基脑苷脂可进一步逐个增加糖基,形成含糖链的鞘糖脂。每一步糖基化反应都在一定的糖基转移酶的催化下进行。这类酶的特异性很强,即对其两个作用物,单糖供体及糖基接受体,具有双重特异性。这些酶都是膜结合的或嵌入膜中的,并可能以复合酶体的形式存在。每一种多酶复合体合成一种糖脂。前体一旦结合到复合酶体便按预定顺序进行连续糖基化,直至生成特定的糖链。糖基化的单糖供体为糖核苷酸。糖脂的硫酸化常发生在Gal基第3位碳原子上,由3'-磷酸腺苷5'-磷酰硫酸(PAPS)提供硫酸,形成硫酸酯键。
鞘糖脂的分解代谢在溶酶体水解酶的催化下进行。糖链的降解由各种外切糖苷酶催化,从非还原(游离)末端逐个切下糖基。神经节苷脂的唾液酸基虽多在糖链的非还原末端,然而仅在无其他单糖作为末端基的情况下才能被唾液酸酶(又称神经氨酸酶)催化水解。糖链完全水解后产生的神经酰胺有两条去路:或作为合成的原料被利用,或进一步在神经酰胺酶的催化下水解为脂肪酸及鞘氨醇。硫酸鞘糖脂的分解还需要硫酸酯酶参加。
生物学作用甘油糖脂主要存在于植物的光合组织及微生物的质膜中。在哺乳动物仅见于神经组织、睾丸和精细胞。生物学作用尚不详。微生物质膜中的甘油糖脂结构差别很大,在微生物分类上有一定价值。植物中的甘油糖脂与叶绿素的构造及光合作用有关。
动物的神经组织中糖脂含量丰富。在中枢神经系统,脑苷脂(主要是半乳糖脑苷脂)的含量在脑白质中很高,此外鞘氨醇半乳糖苷及硫酸化半乳糖脑苷脂分布亦很普遍,并有痕量甘油糖脂。神经节苷脂在脑灰质中含量较高。脑苷脂和硫酸脑苷脂还与鞘磷脂共同构成神经髓鞘的脂类。其合成不足或代谢障碍可严重影响髓鞘的发生和形成,破坏髓鞘的正常结构,引起病变。
鞘糖脂作为各种细胞质膜及细胞内膜不可缺少的成分,含量虽少而生物学作用相当复杂。糖脂作为膜的结构成分可改变膜的物理性质,如赋予质膜一定的刚性,使膜处于最适于发挥其作用的物理状态。由于糖脂具有抗原性,质膜中的糖脂可作为表面抗原。例如,ABO(H)、Ii、Lea/ Leb等血型抗原决定簇不仅存在于红细胞及其他非红细胞质膜的糖蛋白糖链的末端,亦存在于质膜鞘糖脂糖链的末端。在同一组织中合成的糖蛋白及糖脂的寡糖链,其核心部分(还原末端)虽不相同,而末端部分(游离的非还原末端)却极其相似,因此可以共有某些功能。发生恶性转化的细胞表面常出现肿瘤相关的糖脂抗原。例如,人、小鼠及仓鼠的黑素瘤细胞有共同的表面抗原CM3,而正常黑素细胞及其他肿瘤细胞基本不具有这种表面抗原。另外,在各种自发及诱发的人及鼠类肝癌细胞表面,GD3含量大幅度增高,并与肿瘤的发展相平行。制备肿瘤相关抗原的抗体对肿瘤的诊断及治疗有一定意义。鞘糖脂还可作为细胞表面标志,对研究和认识免疫细胞、胚胎发育、细胞分化及肿瘤细胞很有意义。例如,人胸腺细胞可按其是否与识别 α半乳糖的花生凝集素 (PNA)相结合而分为PNA+PNA-两个亚群。正常人周缘血中PNA+淋巴细 胞甚少(1~2%),急性淋巴性白血病及髓性白血病患者有半数出现PNA+细胞升高(≥20%);而慢性白血病及急性白血病的缓解期很少见PNA+淋巴细胞增高。因此测定周缘血中PNA+淋巴细胞的百分率有助于预后的判断。质膜中的鞘糖脂与细胞表面生物活性分子,如受体及酶的关系,更是十分复杂。有的可能就是受体或受体复合物的一个组成部分,有的则为受体糖蛋白构象的维持者或功能的调节者。例如,GM1可作为霍乱毒素的受体,可高效、特异地中和霍乱毒素,抑制霍乱毒素的特异性毒性作用。具有 GM1寡糖结构的膜糖蛋白亦具有霍乱毒素受体的作用。此外,GD1b、GT1b可与破伤风毒素结合,并使之丧失毒性。破伤风毒素属于神经毒。突触膜中的上述神经节苷脂作为破伤风毒素的受体可能是产生破伤风神经症状的原因。巨噬细胞移动抑制因子 (MIF)的受体是一种岩藻糖化的脑苷脂。一些含糖肽类激素,如促甲状腺素、黄体化激素、绒毛膜促性腺激素等,与其靶细胞的结合可被某些神经节苷脂抑制。干扰素的抗病毒活性可被神经节苷脂增强,本来对干扰素无反应的细胞经与神经节苷脂作用(外源性神经节苷脂嵌入于质膜中)后可变为有反应性。一些细胞外基质的细胞表面受体虽已鉴定为糖蛋白,但对一定的糖脂具有依赖性。它们在细胞表面共分布,在体外共纯化,去除糖脂则受体功能丧失。某些病毒亦与一定的鞘糖脂有亲和性。以上种种事实都表明糖脂与多种生物活性分子乃至微生物的受体有密切关系。另外,位于神经末梢及突触前膜的神经节苷脂及硫酸鞘糖脂,由于其所带负电荷可与带正电荷的乙酰胆碱、5-羟色胺、去甲肾上腺素及组胺等神经递质结合,从而有助于神经冲动的传导。除受体外,膜中酶的结构及功能亦与神经节苷脂有密切关系。例如,神经节苷脂GM1可活化Na+,K+-ATP酶,从而调节细胞膜的离子泵。腺苷酸环化酶及3',5'-环核苷酶磷酸二酯酶等亦可被一些神经节苷脂活化。此外,有一些膜蛋白并无疏水肽段,因此不能直接嵌入于膜脂双层,它们借助于膜中的磷脂酰肌醇糖脂而“锚”定于膜上。例如乙酰胆碱脂酶、碱性磷酸酶、5'-核苷酸酶、氨基肽酶P以及脂蛋白脂肪酶等都以其羧基末端的氨基酸残基与磷脂酰肌醇糖脂糖链末端借氨基乙醇共价结合。此外,脑及淋巴细胞表面的Thy-1抗原以及肿瘤细胞的癌胚抗原(CEA)等亦属此例。
鞘糖脂参与细胞间的识别及粘合。例如,Gbos4Cer可能参与神经肌肉接头的识别过程;GM2可能与视网膜顶盖的识别过程有关。硫酸化鞘糖脂还参与细胞与细胞外基质的作用。一些细胞外基质糖蛋白,如层粘连蛋白、血小板反应蛋白及冯・维勒布兰德氏因子等,均可与硫酸化鞘糖脂结合。这种结合不但亲和力高而且具有特异性。
鞘糖脂参与细胞的生长控制。鞘糖脂在质膜中的表达随细胞周期而改变。细胞在体外培养时,细胞密度由于增殖逐渐增加,乃至彼此接触。这时伴有一定种类的鞘糖脂合成增多,其在细胞表面的含量亦增高。这称为“接触延伸反应”。与此同时细胞延长,增殖减慢。加入外源性的相关鞘糖脂,可使旺盛增殖的细胞减慢增殖速度。可见,鞘糖脂合成的“接触延伸反应”可能参与细胞生长的控制。转化细胞及肿瘤细胞鞘糖脂的合成发生改变;某些糖脂的合成发生障碍,同时又可出现在对应的正常细胞很少或没有的鞘糖脂,并有唾液酸化增强现象,还有一些鞘糖脂在细胞表面从隐蔽转变为暴露。恶性细胞表面鞘糖脂的这些异常可能与其生长失控有关。分化诱导剂(丁酸钠及维生素 A酸等)可增加某些糖基转移酶的活性,促进一定的鞘糖脂合成,同时恢复某些转化细胞在生长时的密度抑制(又称接触抑制)。有些神经节苷脂具有生长因子样作用,另一些有生长抑制因子样作用。
细胞表面的鞘糖脂在分化过程中亦发生显著改变。在红细胞和前脑细胞发育过程中以及成肌细胞和小肠上皮细胞分化过程中鞘糖脂的量或质发生变化。小肠上皮隐窝的未分化细胞表面含有乳糖基神经酰胺及葡萄糖脑苷脂,不含神经节苷脂GM3及其合成酶。在12小时内逐渐沿基膜向刷状缘(绒毛顶)迁移,并进行分化,至分化为含消化酶的成熟细胞时上述两种中性鞘糖脂的含量降至很低而GM3及其合成酶的含量显著升高。此外,GM3还可促进肝癌细胞分化。
鞘糖脂贮积病某一种或几种糖苷酶的先天性缺失可导致一定种类的鞘糖脂在神经系统及其他器官中堆积而产生结构及功能障碍,如精神迟钝,运动障碍,肝、脾肿大,眼盲、耳聋等,常致早夭。上表中列出某些鞘糖脂贮积病。这些先天性酶缺失为常染色体隐性遗传(见脂质贮积病)。