[拼音]:jinhua
[外文]:evolution
生物群体的遗传组成部分或全部的不可逆转变。这种转变主要是基于生物与其环境的相互作用。evolution一词来自拉丁文“evolvere”表示展开或把一个卷紧的卷松开的意思。
进化思想的发展
古代的进化思想
关于万物互相转化和演变的自然观可以追溯到人类文明的早期。例如,中国《易经》中的“阴阳”、“八卦”说,把自然界还原为天、地、雷、风、水、火、山、泽八种基本现象,并试图用“阴阳”、“八卦”来解释物质世界复杂变化的规律。古希腊阿那克西曼德(约公元前 6世纪)认为生命最初由海中软泥产生,原始的水生生物经过蜕变(类似昆虫幼虫的蜕皮)而变为陆地生物。这可能是世界上最早的关于生命起源和演变的假说。
特创论和目的论
中世纪的西方,基督教圣经把世界万物描写成上帝的特殊创造物。这就是所谓“特创论”。与特创论相伴随的目的论则认为自然界的安排是有目的性的,“根据这种理论,猫被创造出来是为了吃老鼠,老鼠被创造出来是为了给猫吃,而整个自然界创造出来是为了证明造物主的智慧”(恩格斯:《自然辩证法》·导言)。
转变论与不变论的争论
从15世纪后半叶的“文艺复兴”到18世纪,是近代自然科学形成和发展的时期。这个时期在科学界占统治地位的观点是不变论。当时这种观点被I.牛顿和C.von林奈表达为科学的规律:地球由于所谓“第一推动力”而运转起来,以后就永远不变地运动下去,生物种原来是这样,现在和将来也是这样。到了18世纪下半叶,自然科学的发展不断与这种不变观点相冲突。I.康德的“天体论”首先在不变论自然观上打开了第一个缺口;随后,“转变论”的自然观就在自然科学各领域中逐渐形成。这个时期的一些著名的生物学家,往往在两种思想观点中徬徨。例如林奈晚年在其名著《自然系统》一书中删去了物种不变的词句;法国生物学家G.-L.de布丰虽然把“转变论”带进了生物学,但他一生都在“转变论”和“不变论”之间徘徊。J.-B.de拉马克在1809年出版的《动物哲学》一书中详细阐述了他的生物转变论观点,并且始终没有动摇。
中国自汉、唐以来,随着农业、医药的发展,对生命自然现象的解释也出现过不少关于物种可变的描述和“转变论”的观点。例如北魏贾思勰( 6世纪)在《齐民要术》中,唐郭橐驼在《种树书》中,明末宋应星在《天工开物》中都有关于动植物通过杂交或植物通过嫁接而引起变异的描述。唐颜真卿( 8世纪)、北宋沈括(11世纪)都根据“沧海桑田”的观点对化石的成因作过正确的解释。
均变论与灾变论
英国地质学家C.莱尔在19世纪早期首先将“转变论”带进地质学。他以对现代地壳状态的观察结果来推论古代的地质现象,认为地球经历了缓慢、渐进的变化。这种看法被称为均变论。反之,法国古生物学家G.居维叶则提出灾变论。认为生物的改变是突然的,每次全部消失之后又全部被重新创造出来。他认为地球经历了多次这样的灾变。
活力论和终极目的论或直生论
18世纪末到19世纪后期,大多数动植物学家都没有认真地研究生物进化,而且偏离了古希腊唯物主义传统,坠入唯心主义。这个时期的一些进化理论多半是臆测的,这些理论虽然承认生物种可以转变(所以仍属于“转变论”范畴)但把进化原因归于非物质的内在力量,如“活力”或“活的驱动”等。活力论认为是生物的“内部的力量”即活力驱动着生物的进化,使之越来越复杂完善。但活力论缺乏实际的证据,是一种唯心的臆测。最有名的活力论者就是法国生物学家拉马克。19世纪后期出现的“终极目的论”或“直生论”,认为生物进化有一个既定的路线和方向而不论外界环境如何变化(例如马始终按身体由小变大、齿冠由低变高以及由趾变为蹄的路线发展进化)。古生物学家T.艾默、H.F.奥斯本等是直生论的著名代表。
进化学说
拉马克学说
后人把拉马克对生物进化的看法称为拉马克学说或拉马克主义,其主要观点是:
(1)物种是可变的,物种是由变异的个体组成的群体;
(2)在自然界的生物中存在着由简单到复杂的一系列等级(阶梯),生物本身存在着一种内在的“意志力量”驱动着生物由低的等级向较高的等级发展变化;
(3)生物对环境有巨大的适应能力;环境的变化会引起生物的变化,生物会由此改进其适应;环境的多样化是生物多样化的根本原因;
(4)环境的改变会引起动物习性的改变,习性的改变会使某些器官经常使用而得到发展,另一些器官不使用而退化;在环境影响下所发生的定向变异,即后天获得的性状,能够遗传。如果环境朝一定的方向改变,由于器官的“用进废退”和获得性遗传,微小的变异逐渐积累,终于使生物发生了进化。拉马克学说中的“内在意志”带有唯心论色彩;“后天获得性”则多属于“表型变异”,现代遗传学已证明它是不能遗传的。
达尔文学说
1858年7月1日C.R.达尔文通过20多年的辛勤工作(包括5年的环球考察,20年的资料整理和构思)终于在伦敦林奈学会上宣读了他的关于物种起源的论文。与达尔文同时宣读的另一篇结论相同的论文是由一位研究马来群岛动植物的学者A.R.华莱士提出的。后人称他们的自然选择学说为达尔文-华莱士学说。达尔文在1859年出版的《物种起源》一书中系统地阐述了他的进化学说。其核心自然选择原理的大意如下:生物都有繁殖过剩的倾向,而生存空间和食物是有限的,所以生物必须“为生存而斗争”。在同一种群(群体)中的个体存在着变异,那些具有能适应环境的有利变异的个体将存活下来,并繁殖后代,不具有有利变异的个体就被淘汰。如果自然条件的变化是有方向的,则在历史过程中,经过长期的自然选择,微小的变异就得到积累而成为显著的变异。由此可能导致亚种和新种的形成。
达尔文的进化理论,从生物与环境相互作用的观点出发,认为生物的变异、遗传和自然选择作用能导致生物的适应性改变。它由于有充分的科学事实作根据,所以能经受住时间的考验,百余年来在学术界产生了深远的影响。但达尔文的进化理论还存在着若干明显的弱点。一是他的自然选择原理是建立在当时流行的“融合遗传”假说之上的。按照“融合遗传”的概念,父、母亲体的遗传物质可以象血液那样发生融合;这样任何新产生的变异经过若干世代的融合就会消失,变异又怎能积累、自然选择又怎能发挥作用呢?二是达尔文过分强调了生物进化的渐变性;他深信“自然界无跳跃”,用“中间类型绝灭”和“化石纪录不全”来解释古生物资料所显示的“跳跃性”进化。他的这种观点近年正越来越受到“间断平衡论”者和“新灾变论”者的猛烈批评。
达尔文以后进化论的发展
早在1865年奥地利植物学家G.J.孟德尔就从豌豆的杂交实验中得出了颗粒遗传的正确结论。他证明遗传物质不融合,在繁殖传代的过程中,可以发生分离和重新组合。20世纪初遗传学建立,T.H.摩尔根等人进而建立了染色体遗传学说,全面揭示了遗传的基本规律。这本应弥补达尔文学说的缺陷,有助于进化论的发展;但当时大多数遗传学家(包括摩尔根在内),都反对达尔文的自然选择学说。人们对达尔文进化论的信仰,发生了“严重的危机”。
(1)新拉马克主义与新达尔文主义 在19世纪末到20世纪初这个时期出现过一些新的进化学说。荷兰植物学家H.德·弗里斯在20世纪初根据月见草属的变异情况提出“物种通过突变而产生”的“突变论”,而反对渐变论。这个理论得到当时许多遗传学家的支持。某些拉马克学说的追随者们虽然抛弃了拉马克的“内在意志”概念,但仍强调后天获得性遗传,并认为这是进化的主要因素。50年代在苏联由Т.Д.李森科所标榜的“米丘林学说”强调生物在环境的直接影响下能够定向变异、获得性能够遗传。所有这些观点被称为“新拉马克主义”。A.魏斯曼在1883年用实践来证明“获得性遗传”的错误,强调自然选择是推动生物进化的动力,他的看法被后人称为“新达尔文主义”。
(2)现代综合进化学说 20世纪20~30年代首先由R.A.费希尔、S.赖特和J.B.S.霍尔丹等人将生物统计学与孟德尔的颗粒遗传理论相结合,重新解释了达尔文的自然选择学说,形成了群体遗传学。以后C.C.切特韦里科夫、T.多布然斯基、J.赫胥黎、E.迈尔、F.J.阿亚拉、G.L.斯特宾斯、G.G.辛普森和J.W.瓦伦丁等人又根据染色体遗传学说、群体遗传学、物种的概念以及古生物学和分子生物学的许多学科知识,发展了达尔文学说,建立了现代综合进化论。现代综合进化论彻底否定获得性状的遗传,强调进化的渐进性,认为进化是群体而不是个体的现象,并重新肯定了自然选择的压倒一切的重要性,继承和发展了达尔文进化学说。
(3)中性学说和间断平衡论 1968年,日本学者木村资生根据分子生物学的材料提出了“中性突变-随机漂变假说”(简称中性学说)。认为在分子水平上,大多数进化改变和物种内的大多数变异,不是由自然选择引起的,而是通过那些选择上中性或近乎中性的突变等位基因的随机漂变引起的,反对“现代综合进化论”的自然选择万能论观点(见分子进化的中性学说)。
1972年N.埃尔德雷奇和S.J.古尔德共同提出“间断平衡”的进化模式来解释古生物进化中的明显的不连续性和跳跃性,认为基于自然选择作用的种以下的渐进进化模式,即线系渐变模式,不能解释种以上的分类单元的起源,反对“现代达尔文主义”的“唯渐进进化”观点。目前争论仍在继续中(见间断平衡论)。
小进化与大进化
早在40年代,美国遗传学家R.B.戈德施米特就认为,通常的自然选择,只能在物种的范围内,作用于基因而产生小的进化改变,即小进化;而“进化中的决定性步骤、走向大进化的第一步、由一个种变为另一个种的进化步骤则需要另一种进化方式,而不是靠微小突变的积累”。戈德施米特所提出的“大进化”方式,就是通过他所假设的“系统突变”(涉及整个染色体组的遗传突变)而产生所谓“有希望的怪物”,可能偶然遇到合适的小生境,从而能立足并繁衍下去。这样就可以一下子产生出一个新种、甚至一个新属或新科。
后来,美国古生物学家G.G.辛普森同意把进化的研究分成两大领域:研究种以下的进化改变的“小进化”和研究种以上层次的进化的“大进化”。辛普森虽然接受了戈德施米特的“小进化”与“大进化”的名称,但并不同意戈氏的观点,他并不认为“小进化”与“大进化”是各自不同的或彼此无关的进化方式。在辛普森看来,生物学家主要是研究种以下的现生生物群体的小尺度的进化,即小进化,而古生物学家则主要以化石为对象研究种以上的分类单元在地质时间尺度上的进化,即大进化。后来的学者有的接受戈德施米特的概念,有的则赞同辛普森的看法,还有的则将两种概念调和起来。
小进化研究种以下的进化改变,包括:
(1)小进化的因素和机制,研究遗传突变、自然选择、随机现象(如遗传漂变)等因素如何在个体和群体的层次上起作用,以及如何引起群体的遗传组成的改变等。
(2)种形成,研究新种的形成方式和过程,研究小进化因素如何导致同种的群体之间的隔离的形成和发展,研究种内分化和由亚种、半种到完全的种的发展过程等。
大进化研究种以上的分类单元在地质时间尺度上的进化改变,其对象主要是化石,最小研究单位是种。主要研究内容包括:
(1)种及种以上分类单元的起源和大进化的因素;
(2)进化型式,在时间向度上进化的线系的变化和形态;
(3)进化速度,形态改变的速度和分类单元的产生或绝灭速度,种的寿命等;
(4)进化的方向和趋势;
(5)绝灭的规律、原因及其与进化趋势、速度的关系等。
小进化与大进化在物种这一层次上相互衔接,事实上小进化与大进化都研究物种形成。关于小进化与大进化的关系问题,近年学术界展开了激烈的争论。间断平衡学派认为不能以小进化的机制来解释大进化的事实;而现代综合进化论则认为小进化是大进化的基础,小进化的机制在一定程度上是可以说明大进化的现象的。
进化型式
从进化的观点看,同时生存的不同生物种在时间的向度上可以回溯到一个共同祖先。因此,按照祖裔关系可以将现时生存的和曾经生存过的生物相互联结起来,这种表示祖裔关系的生物进化系统称为种系发生。生物的种系发生可以形象地表示为一颗树:如果从树根到树顶代表时间向度,主干代表共同祖先,大小分枝代表相互关联的进化线系,这就构成所谓种系发生树或进化树。所谓进化型式就是进化在时间与空间上的特征,也就是种系发生的特征,具体表现在进化树的形态上:枝干的延续和分枝方式、树干的倾斜方向和在空间上的配置、树干的中断等等,它代表着种系发生中线系进化、种形成、绝灭等方面的特征。
进化树或种系发生树是研究者根据各方面的资料综合推理而构想出来的,由于研究者所掌握的资料不同、观点不同、构造出来的进化树的形态也不尽相同。因此,关于进化型式常常有不同观点的争论。种系发生是进化生物学家与分类学家共同关心的,古生物学家和生物分类学家对进化型式的研究贡献最多。
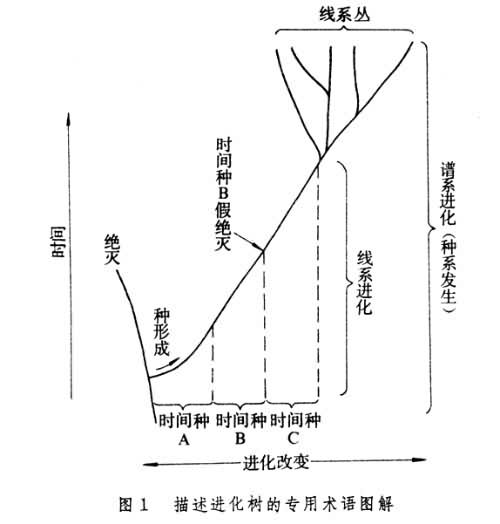
描述进化树的专用术语
如果以时间(通常是地质时间)为纵坐标,进化改变为横坐标,则在这个坐标系中一个瞬时存在的生物相当于一个点,一个世代延续的种则在坐标系中相当于一条由下向上延伸的线,这个代表种在时间向度上延续的线就叫线系。在一个线系之内发生的进化改变称为线系进化,表现为线系在“时间-进化改变”坐标系中朝某一方向倾斜,倾斜度代表该线系的进化速度。如果一个种由线系进化所造成的形态学改变足以构成不同于原来种的分类单元,在古生物学上称为时间种。古生物学家往往为了使化石分类与相应的地层划分相配合而把一个线系由下而上地划分出若干个相继的、渐变的时间种。一个线系在某一地质时间终止,叫做绝灭,它意味着一个种的延续中断,也就是没有留下直接的后裔。在一个线系之内,由一个时间种过渡为相继的另一时间种时,前一个时间种的终止并不意味着真正的延续中断或绝灭,在古生学上称为假绝灭。假绝灭是人为的。在一个生物种之内分化出新种从而造成线系的分枝,叫做种形成。关于种形成的定义存在着不同的意见,有些学者认为种形成等于线系分枝,另一些学者认为种形成不一定伴随线系分枝。由同一条线系分枝出来的若干线系合起来称为线系丛。通常所说的谱系进化或种系发生就是指若干相关的线系的进化,线系进化则指一个线系之内的进化(图1)。
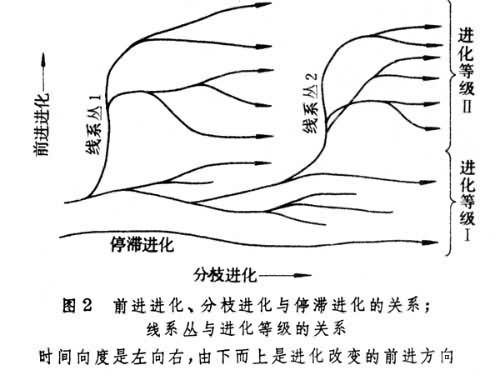
在谱系进化中有两种性质不同的进化改变。一是形态结构及其功能由简单、相对不完善到复杂和相对完善的前进性(进步的)改变,称之为前进进化;前进进化的结果是造成生物的等级从低级到高级。另一种进化改变是线系分枝,叫做分枝进化;分枝进化的结果是产生新的分类单元和生物歧异度的增长。广义的前进进化包括除分枝进化以外的各种进化改变,既包括前进的(进步的)进化改变,也包括非前进的甚至退化的改变。既无前进进化,又无分枝进化的情况称为停滞进化,如“活化石”之类的情况(图2)。
适应辐射
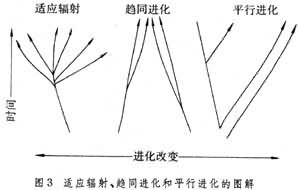
在相对较短的地质时间内,从一个线系分枝出许多歧异的分类单元,叫做辐射。由于辐射分枝产生的新分类群通常是向不同的方向适应进化的(新分类群之间有较大的歧异),所以又称为适应辐射。在种系发生图上适应辐射表现为密集的线系丛(图6)。在分类学上,一个线系丛往往代表种以上的分类单元。可以说:种以上的分类单元往往是通过适应辐射而“快速”产生的。例如:
(1)后生无脊椎动物最早的适应辐射发生在前寒武纪之末,大约6.5~7亿年前;多种多样的无骨骼的软躯体动物印痕化石在世界各大陆的若干地方差不多同一时期的地层中都有发现,其中以澳大利亚的伊迪卡拉动物群为最典型。
(2)在寒武纪一开始,许多有外骨骼的无脊椎动物化石(包括现生的无脊椎动物各门的祖先),在世界各地差不多同时(5.7~6亿年前)的地层中出现。由于众多的门类在较短的地质时间内(2000~3000万年期间)“突然”产生出来,有的古生物学家(如古尔德)称之为“寒武爆发”。
(3)寒武纪以后的无脊椎动物又经历过几次规模较小的适应辐射。例如在早寒武世三叶虫第一次出现后的1500~3000万年间,一共有30个科的三叶虫产生出来;二叠纪末菊石几乎全部绝灭后,到了三叠纪初期的700~800万年间又产生出许多新科;在侏罗纪早期2000万年间产生出六射珊瑚的6个超科。
(4)在早白垩世大约 1000万年间产生出显花植物的许多科。
(5)大部分哺乳动物的“目”是在白垩纪末到第三纪初很快产生出来的。
趋同进化与平(併)行进化
不同的线系各自独立地进化出相似的特征叫做趋同进化,简称趋同。形态结构的进化趋同往往是由于功能的相似,而功能的相似又往往是由于适应于相似的环境。趋同进化的结果是产生功能相同或相似的形态结构,在比较解剖学上叫做同功。与此相对,由于有共同祖先而造成的形态结构的相似叫做同源。例如鸟、蝙蝠和昆虫的翅属于同功器官,它们祖先各自独立地进化出适应于飞翔的翅,这些翅可以说是趋同进化的结果。人的手、鲸的兿、鸟的翅和马的前肢等则属同源器官,是共同祖先经过长期适应辐射的结果。
平(併)行进化(简称平行)是指两个或多个有共同祖先的线系,在其祖先遗传的基础上分别独立进化出相似的特征。通常平行与趋同不易区分,一般说来平行进化既涉及同功又涉及同源;假若后裔间的相似程度大于各自的祖先之间的相似程度则可称为趋同,若后裔之间的相似程度与其祖先之间的相似程度差不多则可称为平行(图3)。
大进化的模式
影响进化型式(进化树的形态)的两个最主要的因素是形态进化速度和种形成(线系分枝)。进化的改变(指形态学的改变)也相应地包含两个分量,即由线系进化产生的线系进化分量和由线系分枝产生的种形成分量。代表两种对立观点的两种大进化模式之间的主要差别就在于分别强调两个分量中的一方面。
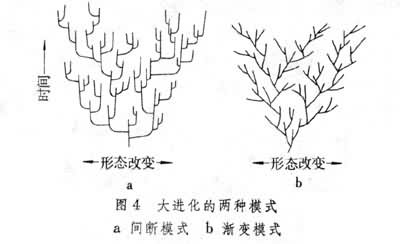
(1)渐变模式:认为形态进化速度多少是恒定的、匀速的,形态进化是渐近的;形态改变主要是线系进化造成的,大多发生在种的生存期间,与种形成无关,种形成(线系分枝)本身只是增加新的进化方向;
(2)间断模式:认为形态进化速度是不恒定、非匀速的,快速的“跳跃”与长期的停顿相交替,即在种形成期间进化加速,种形成后保持相对的稳定。进化的形态改变与种形成相关联,即大部分形态改变是发生在相对较短的种形成时期,在可能长达数百万年的种生存期内不会发生显著的形态改变(图4)。间断模式或间断平衡论是N.埃尔德雷奇和S.J.古尔德于1972年正式提出的(N.埃尔德雷奇和古尔德,1972),之所以称之为“间断平衡”是为了强调大进化的“间断”的特征,即进化在种形成时加速,种形成后又保持稳定的平衡。
渐变论是达尔文以来多数进化学者(包括现代综合论者)的传统观点。现代综合论的先驱者之一J.S.赫胥黎认为,自然选择造成的进化一般说来并不随种形式而加速,大多数种形成是偶然事件,线系进化才是进化的主流。这是典型的线系渐变论观点。间断论的一些基本观点(如快速的种形成,进化的“跳跃”等)可以追溯到20世纪早期。H.德·弗里斯,1905年引用物理学家W.T.开尔文所计算的地球年龄(不超过2000~4000万年)而推论说,种形成不可能象达尔文所描述的那样缓慢,在如此短的时间(当然开尔文的计算是错误的,现代同位素方法测得的地球年龄是46.5亿年)中,种形成必定是大的突变。戈德施米特提出的高级分类单元可以通过染色体“系统突变”而迅速产生的看法也接近于现代间断论的观点。甚至现代综合论的先驱者也都不同程度地强调过种形成过程的“跳跃”特征。例如赖特(1940)认为大物种的小群体进化速率特别高,有可能形成新种;辛普森提出的“量子进化”,也强调了种形成的跳跃形式。E.迈尔(1954)是最早提出“间断进化”观点的,他认为当新种从老种的一个隔离的小群体中产生出来时,常常伴随着快速的进化歧异。他还用这个观点解释了化石记录所显示的进化“鸿沟”。快速的形态进化继之以长时期的形态稳定,这样的实例见之于化石记录;但也有不少实例表明形态进化似乎是在一个线系内发生的。究竟哪一种模式是正确的,目前还不能作出判断,对同样的资料(例如同样的化石记录)往往可以有不同的解释。
总体说来,渐变论和间断论在解释大进化的过程时所强调的方面不同。渐变论或渐变模式强调进化过程的连续和匀速,强调线系进化对总的形态进化改变量的贡献,强调种内的小进化过程中变异的积累,主张自然选择是进化的主要因素。而间断平衡论或间断平衡模式则几乎相反:强调进化过程的不匀速,强调种形成事件对形态进化改变量的贡献,强调小群体内的随机事件、强调大的突变、强调物种选择或环境突变对大进化的作用。双方各有一些有利于自己的证据,当然这些证据本身还需进一步研究才能证实。
有利于间断平衡论的证据有:
(1)古生物的高级分类阶元在相对较短的地质时间内快速出现(同时伴随高级分类阶元的快速绝灭)。在前寒武纪末到早寒武纪的无脊椎动物高级分类群的“快速”出现被形容为“寒武爆发”;在早寒武世的1500~2000万年期间产生了三叶虫的30个科;在二叠纪末菊石大量绝灭之后,到三叠纪的最初800万年期间又从少数物种中产生出好几个新科;侏罗纪初的2000万年期间产生了六射珊瑚6个超科;哺乳类的许多目是在中生代末、新生代初的较短时间内出现的;根据化石记录测算的某些古生物门类的进化速度是不均匀的,在适应辐射期间进化加速。
(2)某些现代生物快速种形成。以东非维多利亚盆地湖泊区的丽鱼科的“爆发式的种形成”为例,据研究维多利亚盆地的众多湖泊形成于75~180万年前,在这些湖泊中生存着500种以上的丽鱼,种数占该科的一半以上。这些丽鱼分布于各个小的湖盆地,地方化很强,每个湖盆也有一个特有的物种群。学者们认为这一地区的500种以上的丽鱼是在75~180万年期间由少数丽鱼祖先快速辐射进化而产生的。这说明,虽然在一般情况下形态学进化速度和分枝进化速度都很缓慢,但在某些合适的环境条件下,进化速度很高。
(3)某些形态结构的进化不可能有中间过渡阶段,只能是跳跃的。例如腹足类的内脏的扭转不可能有中间形态,因为任何中间形态都不适应生存。内脏要么经过 180°扭转而使鳃和肛门等转向头部(前鳃亚纲),要么再经过180°反扭转而使鳃和肛门回到后部(后鳃亚纲)。
(4)某些遗传学的证据支持高级分类单元的跳跃式地产生的假说。例如高等植物的染色体基数往往在较高级的分类群中是恒定的。在木本植物中同一科的植物种往往有相同的染色体基数。草本植物中同一属的植物往往有相同的染色体基数。因此,学者们认为,植物的染色体基数的改变造成一个新属或一个新科,这无疑是进化跳跃。
与此相反,另外一些事实支持渐变论或渐变进化模式:
(1)在自然界中有不完全种(亚种、半种)以及其他有种内分异的复合种(即包含有族、亚种等种下单元的种)的存在说明种形成是通过若干中间阶段的渐进过程。
(2)协进化,在生态上密切相关的(或相互依赖的)物种(例如昆虫或蜂鸟与植物的花,寄生的动物与寄主等)的协同进化不可能采取跳跃的形式。例如蜂鸟的喙的长度形状与其采集花蜜的花的形态结构极为吻合。任何一方的突然的、大的变异都不利于双方。
(3)复杂器官(例如眼)的进化涉及许多相关的结构与机能的协调,因而也不大可能采取跳跃的形式。
(4)重新测算线系进化速度后得出结论认为线系进化速度足够大,可以解释大进化的高级分类单元的快速产生。
进化速率
单位时间内生物进化改变量。衡量进化速率必须确定两个尺度:时间尺度和进化改变量的尺度。时间尺度有两种,即绝对地质时间(根据同位素方法测定的地质时间)和相对地质时间(由于地质年代单位并非等时距,因此在可能的条件下应使用绝对时间尺度;只在不得已时,才使用相对时间尺度)。
进化改变量的衡量比较复杂,通常也应用两种尺度:
(1)形态学尺度,即以形态改变量为衡量尺度。通常用的是体积、长、宽、角度等可测量的数量性状的测量值的变化。
(2)分类学尺度,即以分类单元(种、属、科等)产生的数目为衡量尺度。
相应于上述两种衡量尺度而有两种不同的进化速率:
(1)形态学进化速率──单位时间内形态改变量;形态改变量/时间(百万年)。通常以百万年为时间单位,形态改变量可以直接用测量的单位,也可以将不同的测量单位换算为某种统一的单位。
(2)分类学进化速率──单位时间内新分类单元(种、属、科等)产生的数目:新分类单元产生数目/单位时间(百万年),新分类单元产生的情况比较复杂。新分类单元既包括线系进化产生的时间种,也包括分枝种形成产生的新分类单元。因此,又可以区分出两种情况和两种分类学进化速率:假定所研究的化石材料可以证明是同一线系的,则可计算出某一地质时间内的线系进化速率,即单位时间内时间种的产生数目:线系进化速率=时间种数目/时间(百万年)。如果在时间种的分类上有大致相同的形态学标准,则线系进化速率相当于线系内的形态学进化速率。假定研究的化石材料属于相关的不同线系,则可以计算出某一地质时间内种形成的速率,或分支进化速率:种形成速率=分枝(种形成)事件数目/时间(百万年),不同观点的研究者在研究进化速率时,往往有所侧重。渐变论者常侧重于线系进化速率的研究。但要获得同一线系的完整的化石材料是不容易的。有时因材料不能确证为同一线系,其研究成果也受怀疑。间断论者则更重视分枝种形成速率的研究。
线系进化速率的测定
通常用“时间种”的寿命来测定线系进化速率。“时间种”可以看作是线系进化的片段。因为时间种代表综合的形态变量的测定,同时在某些情况下“时间种”的寿命是可以测量出来的。所以用“时间种”为进化改变量的单位是合适的。某一“时间种”在线系进化中的形态改变达到另一个“新时间种”之前所持续的那段时间就是该“时间种”的寿命。“时间种”的寿命是可以测量的。G.G.辛普森在研究上新世到更新世的哺乳动物进化速度时曾用所谓“分类学生存曲线”的方法。由于“时间种”的寿命反映了线系进化速率(时间种寿命愈短,线系进化速率越高),所以我们可以用“时间种”寿命的倒数作为线系进化速率的测量:
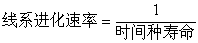
例如一个寿命为 200万年的时间种的线系进化速率可以表示为 0.5“时间种”/百万年,即每百万年产生0.5个“时间种”。如果以大致相同的形态学标准来划分“时间种”,则以“时间种”寿命的倒数作为线系进化速率的测量是比较准确的。但是用这样的方法所测定的某些动物化石的线系进化速率都比较低,例如欧洲上新世至更新世哺乳动物平均线系进化速率为 1.1“时间种”/百万年,极少数达到4“时间种”/百万年,只有1%的线系达到 2.9“时间种”/百万年。用类似的方法测出的人类线系进化速率比哺乳动物平均进化速率还低。海洋无脊椎动物线系进化速率的测量结果数值也很低,例如腕足类的准小石燕贝科中的大多数种的寿命都超过了百万年,线系进化速率在0.3~0.4“时间种”/百万年以下。以这样的低的线系进化速率很难解释大进化,例如欧洲整个更新世的180万年期间至少产生了 15个哺乳动物新属,只有 100个线系可以作为产生新属的亲本,按照1.1“时间种”/百万年的平均速率要在180万年内产生15个新属是不可能的。
然而,正如某些研究者指出的,由于所用的时间间距长,产生很大的误差(由于进化速度不均匀、正向与逆向进化相互抵消等,时间间距愈长,误差愈大,测出的速率愈低),因此,依据化石资料的线系进化速率的测量,仍然存在许多问题。
种形成速率与绝灭速率的测定
如果承认快速进化发生于种形成(线系分枝)期间,或者说种形成是进化加速的位点,那么测定种形成速率是了解进化特征的重要手段。绝灭是进化的另一个侧面,而且种形成速率与绝灭速率相互关联。种形成速率的测定可通过下面的步骤:首先求出物种净增率(R),其次求出绝灭速率(E),最后求出种形成速率(S)。三者的关系是:
S=R+E
物种净增率──种形成过程中物种数目的增长是指数增长,这和生物群体的个体数目增长相似。在测定物种数目的增长时要考虑初始物种数目(基数),换句话说物种净增率是单位时间内物种数目的相对增长。用公式表示则为:
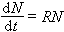
公式中N 代表物种数目,t代表时间,R 就是物种净增率,将上式积分则得:
N=N 0eRt公式中N 0代表初始物种数目,e是自然对数的底。如果已知初始物种数N0和终末物种数N、以及时间t,则可求出R。可将上式变化,得:
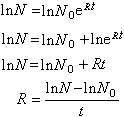
R的单位是“百万年-1”。
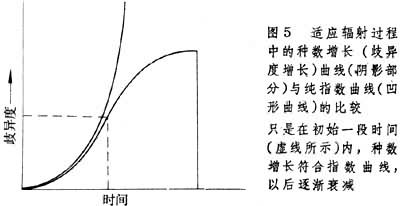
对于不同分类群的 R值的计算结果表明各个大分类群的 R值很不相同,而同一大分类群中的成员具有相近的 R值。实际上种形成过程中物种数目的增长(歧异度的增长)不可能一直遵循指数增长方式。适应辐射期间的物种数目的增长曲线与纯指数增长曲线不一样,前者是“衰变”式的;开始是指数增长,然后增长减缓,曲线向下弯曲(图5)。可以用描述放射性衰变的类似方法来描述适应辐射期间物种数目的增长。但放射性衰变指数是负的,这里是正指数。所以不用“半衰期”,而用种形成“双倍时间”来表示不同分类群的种数目增长特点。当N/N0=2时,
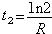
公式中t2就是“双倍时间”,即种数增长一倍所需时间。双倍时间根据R值求出,例如哺乳类平均R值为0.22百万年-1,则双倍时间为t2=ln2/0.22=3.15百万年。
绝灭速率──根据化石资料可以计算出绝灭速率。但通常所计算的绝灭包含了真绝灭和假绝灭。实际上二者不易区分,在一般情况下可以先求出总绝灭速率,假绝灭速率可以估计,三者的关系是
E=E'-e公式中E代表真绝灭速率,E'代表总绝灭速率,e代表假绝灭速率。总绝灭速率可以通过某些间接方法求出。D.M.劳普和W.M.斯坦利曾提出一种方法(1978)其原理是通过测定线系或物种的寿命来求绝灭速率。先假定是简单的情况:设想一组线系,它们的寿命相等,譬如说每个线系都生存500万年。又假定每100万年期间有 5个线系同时存在,每100万年绝灭一个线系(即总数的1/5),它们在时间上均匀分布如图6所示。在这种情况下,绝灭速率可表示为线系寿命的倒数,即
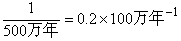
这就是说一个寿命为500万年的线系(或物种),每100万年绝灭 1/5。如果一组线系(或物种)的各成员寿命不等,就可以用适当的方法求出平均寿命和平均绝灭速率。所谓适当方法就是能够尽量消除时距所造成的误差。为此,应以某一瞬时存在的线系或物种的个别绝灭速率来求出总平均绝灭速率,而不能用某一段时间生存的线系或物种来平均。寿命长的物种在某一瞬时存在的概率比寿命短的物种要大,瞬时存在的概率与物种寿命成正比。
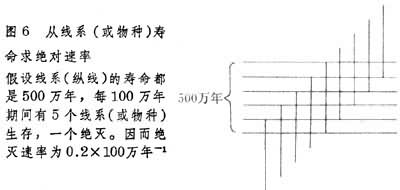
例如,假设在 600万年期间有11个物种生存,其中寿命为 100万年者有6个,200万年者有3个,300万年者有2个(图7a)。由此可以画出在 600万年期间不同寿命的物种的柱状分布图(图7b)。如果我们以 600万年期间生存的11个物种的平均寿命来求平均绝灭速率,显然其结果会偏高,因为我们把寿命短的与寿命长的物种同等看待了。更好的方法是求出每一瞬时(图7中的T)存在的物种数目。求法是将不同寿命的物种数目乘以各自的瞬时存在的概率。寿命为 100万年的瞬时存在概率为1/6,寿命为200万年者为2/6,300万年者为 3/6。这样我们就得出每一瞬时不同寿命的物种的柱状分布图(图7c)。计算结果是不同寿命的物种每一瞬时存在的数目都是 1(6×1/6=1;3×2/6=1;2×3/6=1),也就是说每一瞬时生存的物种总数为3个,其中100万年的1个,200万年1个,300万年的1个。由这3个物种得出的平均寿命为 200万年,平均总绝灭速率为 0.5×100万年-1。如果这里不包含假绝灭速率(假设11个种不包含“时间种”),或假绝灭速率可以估计出来,则可以求出真绝灭速率。
种形成速率(S)由物种净增率(R)与真绝灭速率 (E)之和求得。
进化趋势
从长的时间尺度来看生物进化呈某种方向性,但这并非说自然界存在着既定的进化轨道,这里说的方向性是统计学的趋向。进化是否有方向或是否定向是进化论发展史中争论的主要问题之一。从拉马克的进化梯级说(进化=进步)到直生说都强调进化的定向性。他们认为自然选择控制进化的方向,进化是生物对环境的适应,如果环境变化具有趋向性,则进化也表现出趋向性。随机论者是彻底否定进化定向的。但他们并不反对进化的趋向性,因为随机因素也能造成进化的统计学趋势。因此,进化趋势是任何因素(包括随机的非随机的因素)所造成的进化的统计学趋势。
小进化趋势
小进化趋势是指在生物种的自然群体之内,自然选择作用所造成的表型进化改变的趋向性。这种进化趋势通常是适应局部环境的结果,进化改变发生在线系之内,所以也可称之为线系进化趋势,表现为线系在时间-进化改变坐标系内的倾斜(图8a)。
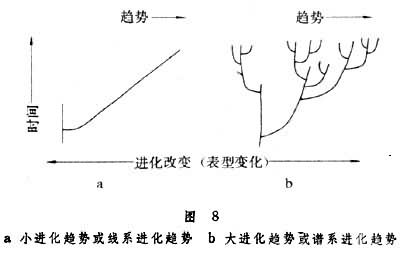
大进化趋势
大进化趋势是指在种系发生中线系分枝的主体的趋向性。大进化趋势发生在种以上的分类单元的谱系进化中,所以又可以称为谱系进化趋势,表现为谱系中的偏斜(图8b )。
(1)造成进化趋势的主要因素:表型分异和分枝分异(或谱系分异)。表型分异指一个单系群在其进化过程中从时间t1到时间t2其平均的表型发生差异或分异(例如,在t2时的平均体积比在t1时的平均体积增大一倍),从而造成谱系的偏斜。造成表型分异的原因有二,一是线系进化(自然选择造成的适应性进化改变),二是种形成或分枝的方向。分枝分异或谱系分异指一个单系群在时间t1到t2之间物种数目的差异性增长,从而造成谱系的偏斜。物种数目的净增长率(R)与种形成速率(S)和绝灭速率(E)有关,即R=S-E。因此,物种数目的差异性增长是由区分性的种形成速率和绝灭速率决定的。假定在某一方向的种形成(线系分枝)速率大于(或小于)另一方向的种形成速率,或某一方向的绝灭速率大于(或小于)另一方向的绝灭速率,就会造成谱系偏斜或谱系进化趋势。
表型分异和谱系分异是构成大进化趋势的两个分量,这两个分量的不同组合可以造成 4种情况:无进化趋势(图9a无表型分异,无谱系分异);进化趋势仅表现为表型分异;进化趋势仅表现为物种净增率(R)的差异(或种形成速率S与绝灭速率的差异);进化趋势表现为表型分异与种净增率差异的结合。

上述 4种情况中在第二种情况下(图9b),左、右侧的物种净增率 R相等,进化趋势表现为线系进化(向右侧倾斜)造成的表型分异。在第三种情况下(图9C),不存在线系进化,但右侧的物种净增率R 大于左侧的,造成谱系主体倾向于右侧。在第四种情况下(图9d),线系进化造成的表型分异和区分性物种净增率相结合,形成右侧的进化趋势。
物种净增率R 的差异可以由种形成速率S 的差异或绝灭速率E 的差异造成。例如,如果一侧的S 大于另一侧则R 值相应地会改变;或是一侧的E 值低于另一侧,则R 值也会不同。
(2)为解释大进化趋势发生的原因不少学者提出过种种假说,但由于缺乏证据,能够成立或有说服力的还很少见。
其一,以小进化趋势(线系进化趋势)解释大进化趋势──某些渐变论者认为进化改变的主要分量是线系进化,大进化的趋势就是由长期稳定的选择压造成的线系进化的趋势。所以大进化趋势与小进化趋势是一致的,造成大进化的原因和机制与造成小进化趋势的相同。例如马的进化,他们认为从始马到现代马是一个长的进化线系。由于长期稳定的选择压力,适应快速奔跑的要求,马的祖先向着增大体躯和改造足趾结构的方向进化。长时期的环境趋向性改变(例如气候的趋向性改变)对生物造成稳定的选择压力可能形成大进化的趋势。当然不是所有的渐变论者都持有这样的观点。例如辛普森就认为进化不是完美的单线系,大进化中包含许多线系分枝,甚至包含与大进化趋势相逆的分枝方向。辛普森认为在马的祖先始马与现代马之间的谱系很复杂,并非一条单线系。
其二,是物种选择说──一些学者认为,自然选择可能在物种层次上起作用,并将小进化机制搬用到大进化中,例如大进化中的物种相当于小进化中的生物个体,大进化中的种形成和绝灭相当于小进化中的个体出生与死亡,大进化中的“种形成”产生的变异相当于小进化中的突变和遗传重组。由于将物种视为自然选择的作用单位,所以称之为物种选择。如果承认自然选择在物种层次上起作用,那么大进化趋势必定是适应趋势,持这种观点的人认为大进化过程中生物体制的复杂化常常包含着适应方面的完善化趋势,生物种之间的竞争可能造成许多进化分枝的区分性绝灭(非随机的绝灭),正是这种非随机因素(区分性绝灭或淘汰)造成大进化的适应趋势,或“进步”趋势。
有些古生物学家认为中寒武世以后三叶虫开始走下坡路,物种分异度下降。从结构上分析,三叶虫与甲壳类相比显然处于劣势:三叶虫缺少强有力的颚、螯和头胸甲,游泳能力也相对较弱,在与甲壳类竞争上很不利。因此,当头足类和鱼类兴盛以后三叶虫大量绝灭,只有一部分留下来并发生了进化改变。这种绝灭并非随机的,那些游泳能力弱、防护能力差的种类绝灭的机会大些。种间竞争、物种选择造成中寒武世以后三叶虫的进化趋势;同时在节肢动物门这个更大的分类群范围内甲壳类又逐步替代了三叶虫类。
个体层次上的自然选择是以随机的遗传变异(突变和重组)和个体间繁殖机会的差异为基础的。物种选择如果存在,其前提应当是种形成事件的随机性和物种的生存或绝灭机会的差异。
种形成事件发生的方向多半是偶然的。例如一个祖先种所遇到的生存环境的性质、环境的综合特征等等多是偶然的,一个种形成事件发生的方向,新种的变异方向是不可预测的。因此,就种形成事件总体而言,也如遗传变异一样具有随机性。化石记录证明某些类群在进化过程中有反复的特点,称之为徘徊进化。例如蚶科动物既有足又有足丝,有的种营固着生活,足丝发达,足退化,但足仍保存于幼体阶段;又有些种在进化中恢复底埋生活,足又发达起来,而足丝仍保留于幼体中。看来种形成事件的方向是既可前进又可后退的。
能够说明大进化趋势是适应的具体证据并不多,但某些古生物学家认为生物进化中结构复杂化和功能完善化的趋势就是适应的。在某一地质时代曾经是成功地适应环境的种类,换了时代就未必成功了,因为又有了更适应的种类出现;中生代的恐龙如果生活于现代未必能占据那么广阔的生境,因为今日有更适应的动物种类;有袋类在澳大利亚是成功的,但竞争不过哺乳类中较晚出现的真兽类。
关于种间竞争导致非随机性绝灭的证据大多是推测性的。例如一些学者认为捕食种(肉食性物种)的适应辐射会导致被捕食种(植食性物种)的绝灭率增高;在食物链的位置愈高,物种之间的竞争愈烈,绝灭率愈高。非随机性绝灭导致区分性的物种净增率 (R),从而引起谱系分异。
其三,谱系飘移说──某些学者,例如D.M.劳普、古尔德(1974)用计算机模拟来证明随机因素对进化谱系趋势的影响。正如在小群体中的遗传漂变能造成小群体对群体总体的大的偏离一样,在小的分枝丛中随机因素(随机的种形成和随机的绝灭)也会造成谱系的显著的偏斜(谱系趋势),这叫做谱系飘移。这方面的实际证据还没有,只是停留在数学方法的推理上。
其四,效应假说──E.S.弗尔巴提出的大进化趋势发生原因的另一种解释是:进化趋势可能是生物种内在的过程和特征的非自然选择的效应,他称之为效应假说。这个假说认为,虽然种形成事件发生的方向(进化改变的方向)是随机的,但不同物种或不同线系可能具有不同的种形成几率;种以下的自然选择造成的适应特征有可能影响或决定不同物种的特有的种形成几率;简言之,不同的物种或线系具有不同的形成新种(分枝)的潜力或可能性。而差异性的种形成就会导致大进化趋势。小进化过程造成的适应性状可能影响种形成速率 S,也间接地影响到绝灭速率E。不同的S与E造成区分性的R,从而造成谱系分异。如广适性的种与狭适性的种在变化的环境条件下,种形成与绝灭速率明显不同。广适性、广布的种在改变了的环境条件下仍能较好地适应,受到的选择压较低,S 和E 增高不显著;反之,狭适性、特化的种在改变了的环境条件下,受到的选择压较高,S 和E 也可能升高。
- 参考书目
- P.L.Forey,The Evolving Biosphere,British Museum (Natural History), Cambridge Univ. Press,London,1981.
- A. Hallam,Patterns of Evolution,Elsevier,Biomedical Press,Amsterdam,1977.
参考文章
- 下列观点不属于达尔文进化理论的是__知识题库
- 研究生物进化最直接、最重要的证据是知识题库
- 下列观点不属于生物进化论的是知识题库
- 在森林古猿进化成人的过程中,下列哪一项是人类的主要标志知识题库
- 木星是太阳系中体积最大的行星,人们对它充满无限退想。为了解木星的形成、进化和结构等,在最新航天科技的支持下,美国于201知识题库
- 爱因斯坦说:“想象比知识更重要,因为知识是有限的,而想象力概括着世界上的一切,推动着进步,并且是知识进化的源泉。严格地讲知识题库
- 下列叙述,属于达尔文进化论的主要观点的是__。①一切生物都经历了由低级向高级,由简单到复杂的发展过程②生存竞争和知识题库
- 浙江进化茶叶是富硒茶 每千克茶叶含硒5.2毫克经济作物
- 迦密山人类进化遗址:梅尔瓦特 河谷/瓦迪艾玛哈尔洞穴群建筑与名胜
- 基因组进化研究促进水稻分子育种改良粮食作物