[拼音]:shengwu shuxue
[外文]:biomathematics
生物学与数学之间的边缘学科。它以数学方法研究和解决生物学问题,并对与生物学有关的数学方法进行理论研究。它的分支学科较多,从生物学的应用去划分,有数量分类学、数量遗传学、数量生态学、数量生理学和生物力学等。这些分支是数学与生物学不同领域相结合的产物,在生物学中有明确的研究范围。从研究使用的数学方法划分,生物数学又可分为生物统计学、生物信息论、生物系统论、生物控制论和生物方程等分支。这些分支与前者不同,它们没有明确的生物学研究对象,只研究那些涉及生物学应用有关的数学方法和理论。
生物数学具有丰富的数学理论基础,包括集合论、概率论、统计数学、对策论、微积分、微分方程、线性代数、矩阵论和拓扑学,还包括一些近代数学分支,如信息论、图论、控制论、系统论和模糊数学等。由于生命现象复杂,从生物学中提出的数学问题往往十分复杂,需要进行大量计算工作。因此,电脑是生物数学产生和发展的基础,成为研究和解决生物学问题的重要工具。然而就整个学科的内容而论,生物数学需要解决和研究的本质方面是生物学问题,数学和电脑仅仅是解决问题的工具和手段。因此,生物数学与其他生物边缘学科一样,通常被归属于生物学而不属于数学。
根据生命科学的需要,生物数学的内容分为以下几个主要方面。
生命现象数量化的方法
所谓生命现象数量化,就是以数量关系描述生命现象。数量化是利用数学工具研究生物学的前提。生物表现性状的数值表示是数量化的一个方面。生物内在的或外表的,个体的或群体的,器官的或细胞的,直到分子水平的各种表现性状,依据性状本身的生物学意义,用适当的数值予以描述。数量化还表现在引进各种定量的生物学概念,并进行定量分析。如体现生物亲缘关系的数值是相似性系数。各种相似性系数的计算方法以及在此基础上的聚类运算构成数量分类学表征分类的主要内容(见数量分类学)。遗传力表示生物性状遗传给后代的能力,对它的计算以及围绕这个概念的定量分析是研究遗传规律的一个重要部分(见数量遗传学)。多样性,在生物地理学和生态学中是研究生物群落结构的一个抽象概念,它从种群组成的复杂和紊乱程度体现群落结构的特点。多样性的定量表示方法基于信息理论。
数量化的实质就是要建立一个集合函数,以函数值来描述有关集合。传统的集合概念认为一个元素属于某集合,非此即彼、界限分明。可是生物界存在着大量界限不明确的、“软”的模糊现象,如此“硬”的集合概念不能贴切地描述这些模糊现象,给生命现象的数量化带来困难。1965年L.A.扎德提出模糊集合概念,模糊集合适合于描述生物学中许多“软”的模糊现象,为生命现象的数量化提供了新的数学工具。以模糊集合为基础的模糊数学已广泛应用于生物数学。
数学模型方法
为了研究的目的而建立,并能够表现和描述真实世界某些现象、特征和状况的数学系统,称为数学模型。数学模型能定量地描述生命物质运动的过程,一个复杂的生物学问题借助数学模型能转变成一个数学问题,通过对数学模型的逻辑推理、求解和运算,就能够获得客观事物的有关结论,达到对生命现象进行研究的目的。
例如描述种群增长最简单的模型是马尔萨斯方程:
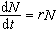
(常数r>0)式中N表示种群的数量;r是种群增长的相对速率。方程的解为
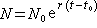
式中N0表示时间为t0时初始种群大小。这个模型简单地描述种群按几何级数增长的过程。
从数学模型获得的结果应该符合实际情况,否则对模型应进行修改,使之尽可能正确地表达生命物质运动的真实情况。模型的不断完善是对生命现象认识逐渐深入的过程。
上述模型的解,种群随时间推后无限增大,这个结果显然不合理。如果考虑有限生存条件的限制,改进之后的模型有费尔许尔斯特-珀尔方程,又称Logistic方程。
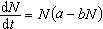
(常数a,b>0)如果初始值取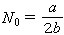 ,方程的解
,方程的解
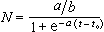
当t →∞,解的渐近值是a/b,它表示种群受生存条件限制不可能超过的极限。这个模型比较正确地表示种群增长的规律,具有广泛用途。
描述捕食与被捕食两个种群相克关系的数学模型是洛特卡-沃尔泰拉方程:
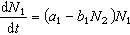
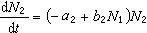
(常数a1、a2、b1和b2>0)其中N1和N2分别表示被捕食和捕食种群的大小。方程的解是
a2lnN1+a1lnN2-b2N1-b1N2=C其中C为积分常数,由初始条件(初始两个种群大小)确定。不同的初始条件得到相应的曲线簇,从曲线的形状可以看出种群此起彼落周期性的变化(图1)。
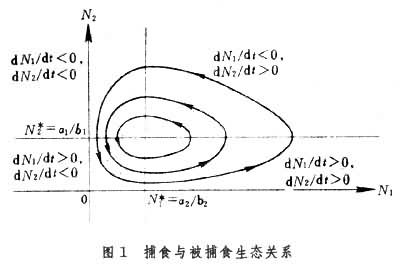
对模型的进一步分析可知,如果捕食与被捕食种群以相同的比例减小,将有利于被捕食种群大量增长。这个结果从理论上说明了不适当地使用农药,在毒杀害虫的同时也杀死了害虫的天敌,而常常导致害虫更猖獗地发生。利用方程的解,还可算出种群变化的近似周期和振幅等十分有意义的结果。
A.L.霍奇金和A.F.赫胥黎从生物膜上电离子的迁移阐明神经兴奋传导的机理。他们建立的模型属于二阶偏微分方程,称霍奇金-赫胥黎方程(H-H方程):
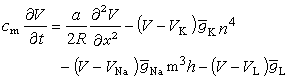
其中V表示神经纤维膜电位,R是轴向电阻率,α是轴突半径,x表示神经纤维轴向距离。等式左边代表膜电容产生的电流分量;右边第一项代表神经纤维横截面电流变化率;右边其余三项分别代表钾、钠和其他离子产生的电流分量。霍奇金曾以枪乌贼神经纤维为实验材料,根据H-H方程计算得到的曲线与实验结果吻合得很好(见生物膜离子通道)。
一种比H-H方程更一般的方程类型,称为反应扩散方程。作为数学模型这一类方程在生物学中广为应用,它与生理学、生态学、群体遗传学、医学中的流行病学和药理学等研究有较密切的关系。60年代,I.普里戈任提出著名的耗散结构理论,以新的观点解释生命现象和生物进化原理,其数学基础亦与反应扩散方程有关。
综合分析方法
由于那些片面的、孤立的、机械的研究方法不能完全满足生物学的需要,因此,在非生命科学中发展起来的数学,在被利用到生物学的研究领域时就需要从事物的多方面,在相互联系的水平上进行全面的研究,需要综合分析的数学方法。
多元分析适应生物学等多元复杂问题的需要、在统计学中分化出来的一个分支领域。它是从统计学的角度进行综合分析的数学方法。多元统计的各种矩阵运算体现多种生物实体与多个性状指标的结合,在相互联系的水平上,综合统计出生命活动的特点和规律性。
生物数学中常用的多元分析方法有回归分析、判别分析、聚类分析、主成分分析和典范分析等。生物学家常常把多种方法结合使用,以期达到更好的综合分析效果。
多元分析不仅对生物学的理论研究有意义,而且由于原始数据直接来自生产实践和科学实验,有很大的实用价值。在农、林业生产中,对品种鉴别、系统分类、情况预测、生产规划以及生态条件的分析等,都可应用多元分析方法。医学方面的应用,多元分析与电脑的结合已经实现对疾病的诊断,帮助医生分析病情,提出治疗方案。
系统论和控制论以系统和控制的观点,进行综合分析的数学方法。
例如有一个生态系统,包括水、一个水生植物种群和一个草食动物种群,研究物质磷在系统中的变化过程。水、水生植物和草食动物含有磷的数量是系统的基本变量,分别以x1、x2和x3表示,称为状态变量;以u表示磷从流水中带进系统的速率,称为输入量;分别以y1和y2表示磷从水中流失和草食动物带出系统的速率,称为输出量。系统内部磷的变化关系见图2。
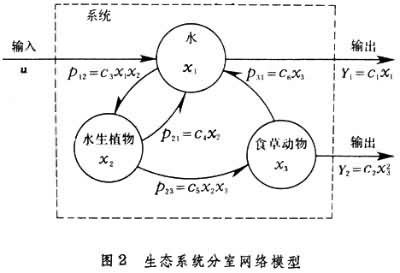
考虑每个状态变量的变化,得到描述该系统的方程,称为状态方程:
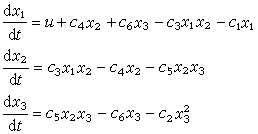
其中Ci(i=1,2,…,6)是一组参数。当参数值、输入、输出以及初始状态给定以后,物质磷在系统中的变化可由方程完全确定。对方程进行分析或者利用电脑求解,就可以认识磷在系统中变化的规律。
实际情况远比这个虚构的例子复杂。一个系统可以是多输入、多输出,状态变量的个数可大到几十,甚至上百,它显示生命活动异常复杂的情形。系统论和控制论的方法没有把那些次要的因素忽略,也没有孤立地看待每一个特性,而是通过状态方程把错综复杂的关系都结合在一起,在综合的水平上进行全面分析。对系统的综合分析也可以就系统的可控性、可观测性和稳定性作出判断,更进一步揭示该系统生命活动的特征。
在系统和控制理论中,综合分析的特点还表现在把输出和状态的变化反馈对系统的影响,即反馈关系也考虑在内。生命活动普遍存在反馈现象,许多生命过程在反馈条件的制约下达到平衡,生命得以维持和延续。对系统的控制常常靠反馈关系来实现。
可控系统的最优控制是控制理论的中心问题。所谓最优控制,就是从实际需要出发设计适当的性能指标,在一定的约束条件下选取输入u(t),使性能指标取最小值。寻求生物系统最优控制的方法常常采用庞特里雅金最小值原理和贝尔曼的动态规划,有关农业、林业、医学和环境问题的最优控制可望获得解决。
概率与统计方法
生命现象常常以大量、重复的形式出现,又受到多种外界环境和内在因素的随机干扰。因此概率论和统计学是研究生物学经常使用的方法。
生物统计学是生物数学发展最早的一个分支,各种统计分析方法已经成为生物学研究工作和生产实践的常规手段(见生物统计)。
概率与统计方法的应用还表现在随机数学模型的研究中。原来数学模型可分为确定模型和随机模型两大类。如果模型中的变量由模型完全确定,这是确定模型;与之相反,变量出现随机性变化不能完全确定,称为随机模型。又根据模型中时间和状态变量取值的连续或离散性,有连续模型和离散模型之分。前述几个微分方程形式的模型都是连续的、确定的数学模型。这种模型不能描述带有随机性的生命现象,它的应用受到限制。因此随机模型成为生物数学不可缺少的部分。
这里举出一种离散的随机数学模型,称为马尔科夫链。考虑具有两个等位基因A与α的群体,如果相应的基因频率分别是p和q,三种基因型AA,Aa和aa在群体中的分配比率构成向量[PHQ](P +H +Q=1)。在一定的假设条件下,按马尔科夫链的数学模型,描述该群本随机交配的遗传过程。经过第一代随机交配,基因型分配比率将从向量[PHQ]转变为
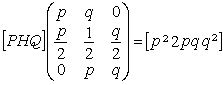
等式左边的矩阵是转移矩阵,不难验证该马尔科夫链是正则的,不动点向量就是[p22pqq2]。
这个结果说明基因频率的不变性,也就是群体遗传学中的哈迪-魏因贝格定律:随机交配的群体在没有外界迁入、定向选择、基因突变和遗传漂变的条件下,基因频率保持不变。
马尔科夫链数学模型不仅对遗传学重要,如果使状态变量代表不同的意义,它还能适用于更广泛的生物学问题,如生态、环境和医学等。下面是一个流行病学的例子。讨论某地区某种传染病的流行,分4个状态:敏感者、患病者、免疫者和死亡。建立的马尔科夫链数学模型可以由转移图的形式表示(图3)。
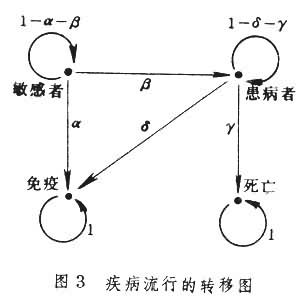
这是一个吸收马尔科夫链,利用这个模型可以分析疾病流行的规律。
不连续的数学方法
不连续性是一切物质存在的基本属性。首先物质和能量两个最基本的概念是不连续的;再看生命现象,物种、个体、细胞、基因等等都是生命活动不连续的最小单位,不连续性表现尤其突出。因此,不连续的数学方法在生物数学中占有重要地位。再举单一种群增长的生态模型讨论。若考虑个体生活年龄,按年龄单位将个体分属于不同年龄组。令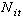 代表在时刻t,年龄为i的个体数;Pi表示年龄在i能活到i+1的存活率;Fi表示年龄在i的增殖率。则新增殖的个体数
代表在时刻t,年龄为i的个体数;Pi表示年龄在i能活到i+1的存活率;Fi表示年龄在i的增殖率。则新增殖的个体数
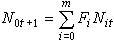
其中m 代表该群体年龄可能达到的上界。于是种群变化的规律可以用下面的矩阵运算表示,
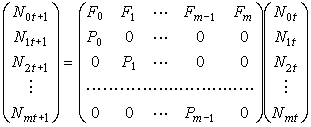
这就是著名的莱斯利模型。这个模型是离散的,它不仅表示种群增长的速度,而且还显示出年龄分布状况,从年龄分布的结构上展示整个种群变化的规律。因而远远胜过前面所举单一种群增长连续模型。
描述生命现象的离散模型有两态和多态之分。马尔科夫链和莱斯利模型都属于多态;两态的模型应生物学的二元表现状态而产生。如神经兴奋沿着神经细胞的轴突,经过突触在阀的控制下传给另一个神经细胞,兴奋波的通过与否就是一个二元表现状态。1943年W.S.麦卡洛克和W.皮茨在布尔代数的基础上,首次给出描述神经传递现象的离散模型。此模型不断改进,并借助电脑加以实现,目前已做到模拟许多较复杂的神经功能,成为探索人类大脑思维奥秘的一个重要手段(见人工智能)。
不连续数学方法还表现在对连续方法的补充。微积分学的基本理论指出,函数的可微性蕴涵着连续性。因此以微分运算为基础的数学模型都是连续的。这些模型只能适用于连续变化范围,对于连续函数出现不连续点或奇点(包括导函数不连续点)情形,将无能为力。而恰恰在这些破坏了连续性的区域,却常常是生物学需要研究的课题。
60年代末,法国数学家R.托姆从拓扑学提出一种几何模型,能够描绘多维不连续现象,他的理论称为突变论。生物学中许多处于飞跃的、临界状态的不连续现象,都能找到相应的跃变类型给予定性的解释。跃变论弥补了连续数学方法的不足之处,现在已成功地应用于生理学、生态学、心理学和组织胚胎学。对神经心理学的研究甚至已经指导医生应用于某些疾病的临床治疗。
继R.托姆之后,跃变论不断地发展。例如E.C.塞曼又提出初级波和二级波的新理论。跃变理论的新发展对生物群落的分布、传染疾病的蔓延、胚胎的发育等生物学问题赋予新的理解。
上述各种生物数学方法的应用,对生物学产生重大影响。20世纪50年代以来,生物学突飞猛进地发展,多种学科向生物学渗透,从不同角度展现生命物质运动的矛盾,数学以定量的形式把这些矛盾的实质体现出来。从而能够使用数学工具进行分析;能够输入电脑进行精确的运算;还能把来自各方面的因素联系在一起,通过综合分析阐明生命活动的机制。总之,数学的介入把生物学的研究从定性的、描述性的水平提高到定量的、精确的、探索规律的高水平。生物数学在农业、林业、医学、环境科学、社会科学和人口控制等方面的应用,已经成为人类从事生产实践的手段。
数学在生物学中的应用,也促使数学向前发展。实际上,系统论、控制论和模糊数学的产生以及统计数学中多元统计的兴起都与生物学的应用有关。从生物数学中提出了许多数学问题,萌发出许多数学发展的生长点,正吸引着许多数学家从事研究。它说明,数学的应用从非生命转向有生命是一次深刻的转变,在生命科学的推动下,数学将获得巨大发展。
当今的生物数学仍处于探索和发展阶段。生物数学的许多方法和理论还很不完善,它的应用虽然取得某些成功,但仍是低水平的、粗略的、甚至是勉强的。许多更复杂的生物学问题至今未能找到相应的数学方法进行研究。因此,生物数学还要从生物学的需要和特点,探求新方法、新手段和新的理论体系,还有待发展和完善。
- 参考书目
- U. N. Bhat, Elements of Applied Stochastic Processes,2nd ed.,John Wiley & Sons,Brisbane,1984.
- M.Braun,et al.,Differential Equation Models,Springer-Verlag,New York,1983.
- E.C.皮耶卢著,卢泽愚译:《数学生态学引论》,科学出版社,北京,1978。