[拼音]:jianshu fenlie
[外文]:meiosis
导致生殖母细胞中染色体数目减半的分裂过程(见彩图)。它是1883年由 E.van贝内登在动物和1888年由E.A.施特拉斯布格在植物最初发现的。在所有进行有性生殖的生物的生活史中,细胞除进行数次有丝分裂外,还要进行一次减数分裂,使其染色体数目由二倍体(2n)变为单倍体(n)。减数分裂发生在配子形成前的某一时期,所以雌雄配子的核都是单倍的。受精后形成的合子又成为二倍的。由于减数分裂,使每种生物代代都能够保持二倍体的染色体数目。在减数分裂过程中非同源染色体重新组合,同源染色体间发生部分交换,结果使配子的遗传基础多样化,使后代对环境条件的变化有更大的适应性。动物的减数分裂发生在形成配子之前。在雄性个体的精细管中,每个精母细胞通过减数分裂形成四个精细胞,精细胞分化成为精子。在雌性个体的卵巢中,卵母细胞通过减数分裂形成卵细胞。被子植物、裸子植物、蕨类和苔藓植物的生活史都由孢子体和配子体世代组成,减数分裂发生在孢子体世代的末尾。在被子植物的雄蕊花药中,小孢子母细胞通过减数分裂形成四个小孢子。小孢子经有丝分裂发育成雄配子体。雌蕊子房中的大孢子母细胞通过减数分裂形成大孢子。大孢子经有丝分裂发育成雌配子体。藻类和菌类进行减数分裂的时期依物种的不同而有多种情况。极端特殊情况见于水绵和衣藻,它们的配子结合(受精)形成合子,合子的第一次分裂就是减数分裂(见有性生殖)。
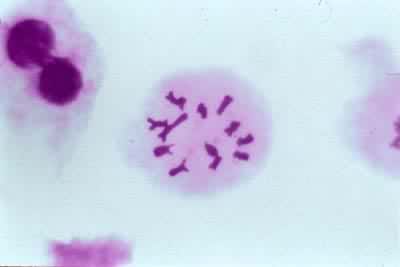
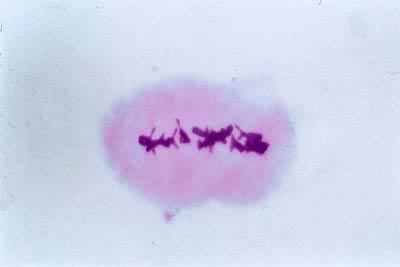
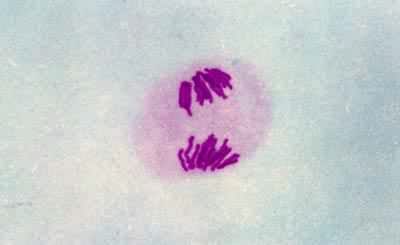
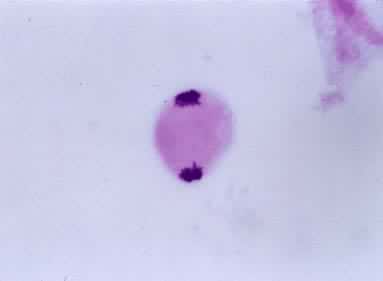
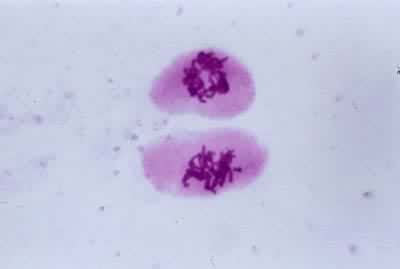
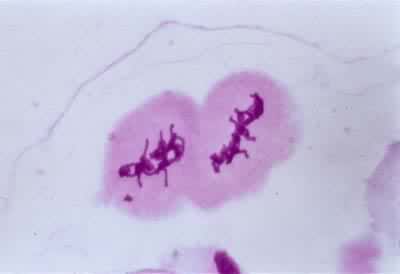
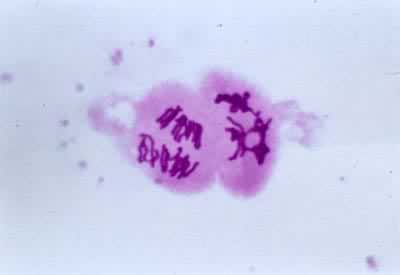
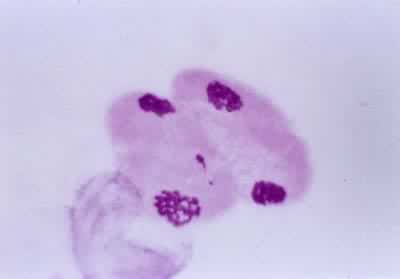
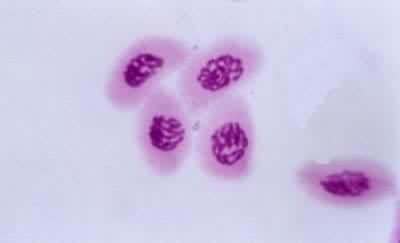
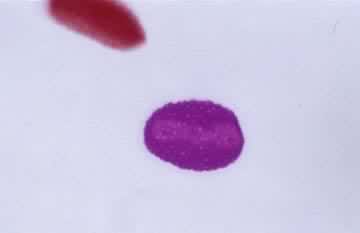
过程
减数分裂是由相继的两次分裂组成的,分别称为减数分裂Ⅰ和减数分裂Ⅱ。在这两次分裂之间一般有一很短的间期,不进行 DNA合成,从而也不发生染色体复制。由于细胞核分裂两次,而染色体只复制一次,所以经过减数分裂染色体数目减半。
减数分裂Ⅰ
前期比较复杂,减数分裂的许多特殊过程都发生在这一时期。因是第一次分裂的前期通常称为前期Ⅰ。它又细分为:
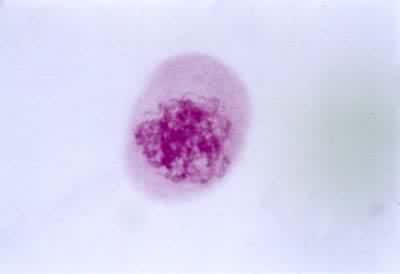
(1)细线期,染色质已集缩成细长的线状结构,每条染色体通过附着板与核膜相连,局部可见念珠状小节称为染色粒。一般认为染色粒是染色线紧密地螺旋折叠的结果。此期核的体积增大,核仁也较大。
(2)合线期亦称偶线期,是同源染色体配对的时期。这种配对称为联会。每对中的两条同源染色体分别来自雌性和雄性生殖细胞,它们在形态和遗传结构上是相似的。同源染色体的配对一般是从靠近核膜的一端开始,有时在染色体全长的若干点上也同时进行联会。配对是靠两条同源染色体间沿长轴形成的联会线复合体实现的。配对后的每对同源染色体称二价体。由于联会,细胞中的染色体由2n条单价体成为n条二价体,虽然DNA含量未变,但数目看起来减少了一半。
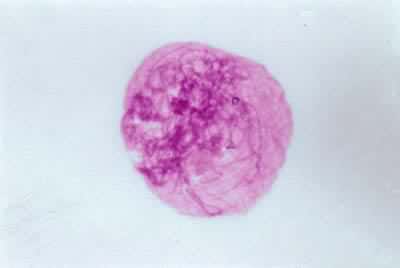
(3)粗线期,染色体明显缩短变粗。因此,在一些染色体数不多的生物,此时可以看出细胞中有几个二价体。联会的两条同源染色体结合紧密,只在局部位置上有时可分辨出是两条染色体。在有些植物,例如玉米,此期二价体的着丝粒、染色粒、异染色质区和核仁组织区都可以看清,因此利用这些特征和染色体长度可做核型分析。在粗线期每条染色体实际已由两条染色单体组成,来自同一染色体的称为姐妹染色单体,对它们来说来自另一条染色体的单体则是非姐妹染色单体。在粗线期同源染色体的非姐妹染色单体间发生局部交换。因为联会非常紧密,这种交换过程不能直接看到。但可根据下一时期(双线期)看到的交叉现象,以及同源染色体间基因重新组合的遗传学事实,判断发生了这种过程。粗线期核仁仍然很大,含有很多RNA。
(4)双线期,联会的两条同源染色体开始分离,但在许多称作交叉的点上它们还连在一起。此期可以看清,联会的两条染色体都分别由两条染色单体组成。故每个二价体由四条染色单体构成。交叉发生在两条非姊妹染色单体之间。一般认为,交叉是发生了交换的结果。双线期的染色体进一步缩短,此时联会丝复合体已消失。在植物一般双线期的时间比细线期、合线期和粗线期都短得多。但人和动物的卵母细胞常长期停留在减数分裂的双线期。例如人的卵母细胞在五个月的胎儿中已达双线期,而一直到排卵时都停在此期。
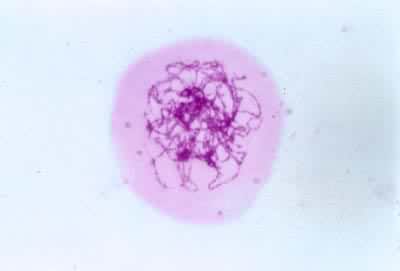
(5)终变期亦称浓缩期,二价体显著收缩变粗,并向核的周边移动,在核内较均匀地分散开。核仁此时开始消失,但有的植物,如玉米、水稻,在终变期的早期核仁仍然很大,并染色很深。交叉数目减少,终变期末有些二价体的同源染色体只在末端连在一起。
中期Ⅰ核膜解体后二价体分散在细胞质中。其中的一条染色体通过牵引丝与一极相连,而另一条染色体则通过牵引丝与另一极相连。最后二价体排列于赤道区,形成赤道板。
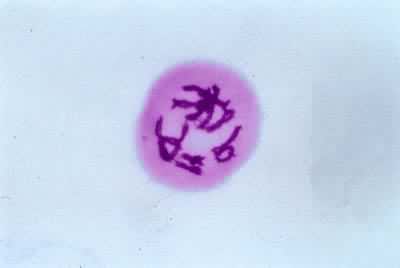
后期Ⅰ每个二价体的两条同源染色体分开,移向两极。n个二价体成为n条单价染色体,此时 DNA含量减半。二价体中那条染色体移向那一极是随机的。
末期Ⅰ染色体各自到达两极后逐渐解螺旋化,变成细线状。核膜重建,核仁重新形成,同时进行细胞质分裂。许多植物在减数分裂Ⅰ只发生核的分裂,而细胞质分裂在减数分裂Ⅱ的末期进行,使四个核同时分开。
间期
在减数分裂Ⅰ和减数分裂Ⅱ之间的间期很短,不进行染色体复制。这时每条染色体已是由两条染色单体构成了。在有些生物甚至没有这个间期,而由末期Ⅰ直接转为前期Ⅱ。
减数分裂Ⅱ
这次分裂基本上与有丝分裂相同。前期Ⅱ时间较短。中期Ⅱ染色体排列于赤道面,两条染色单体的着丝点分别向着两极,形成赤道板。后期Ⅱ时两条染色单体分开,移向两极。到达两极的子染色体为n数,并且每条子染色体只由一条染色单体构成。末期Ⅱ时两极的子染色体解螺旋化。形成核膜,出现核仁,经过细胞质分裂,完成减数分裂过程。新产生的每个细胞都变成了单倍体。
同源染色体的联会
减数分裂机制的核心是同源染色体的联会和交换。用电子显微镜观察发现,在配对的同源染色体之间沿纵轴形成联会线复合体(SC)。靠它把每对同源染色体稳定地联在一起。SC的细微结构在各种动植物中大体相同。两侧是约 400埃宽电子密度很高的侧部组分,很暗。两侧之间为约1000埃的低电子密度的中间区,在电子显微镜下较为明亮。在中间区的当中为中央组分,约 300埃是比较暗的区域。SC的这些部分的厚度依生物种类有些差异。侧部组分的外侧纤维和染色体基本纤维之间连接十分紧密,以至难以区分。在侧部组分和中央组分之间有细纤维相连结,称L-C纤维。L-C纤维直径为70~100埃左右。有人推测,L-C纤维在中间区相遇,形成中央组分。此过程称为锁合。侧部组分主要由蛋白质构成,其中也有少量DNA。L-C纤维和中央组分也主要由蛋白质组成。关于其中是否有 DNA尚无确切的证据。关于SC形成的详细过程还不清楚。有实验说明,细胞中的DNA大约有0.3%是在合线期合成的。这种 DNA称“合线期 DNA”。它并不分布在染色体的某一位置上,而是散在于整个染色体上。如果在细线期或合线期加入DNA合成抑制剂。抑制合线期DNA合成,则同源染色体不能配对,SC不能形成。如果在合线期阻抑蛋白质的合成,也会使同源染色体的配对过程遭到阻碍,停止SC形成。实验表明,细胞中存在着许多基因支配联会过程,有的促进、有的抑制联会。
交换的机制
曾经提出过各种模型来解释交换过程的机制,但至今没有一个被公认。从分子水平理解交换的机理必然包括DNA的断裂和再结合过程。DNA的切断必须有核酸酶参加,在百合中发现有一种脱氧核糖核酸酶(最适pH为 5.2)在合线期活性开始上升,粗线期达最高峰,而在粗线期末消失。DNA的断口再结合要有连接酶参加。有实验表明,由合线期末到粗线期连接酶的活性上升。在许多材料中(百合、小麦、黑麦、蝾螈、小鼠和人)都看到在粗线期有DNA合成。这时合成的DNA称为“粗线期DNA”(P-DNA)。它与合线期 DNA不同,是由非常短的片断构成的。如果抑制P-DNA的合成,则染色体就会发生断裂。因此 P-DNA对保持染色体的完整性是很重要的。推测P-DNA在DNA断裂的再结合过程中对断口的修复可能起一定作用。在DNA酶的作用下,DNA可产生很多断口,但同源的二条染色体在相同的位置上都产生断口的机率是不高的,只有这样的断口有可能发生交换,其他大部分断口将被修复。同源染色体间实现局部交换,必须在空间上非常靠近,否则交换过程是不可想像的。因此联会是实现交换的前提条件。有许多实验证明,抑制联会过程,不形成SC,则交换过程不能发生,也看不到交叉现象。例如,在合线期抑制蛋白质的合成或合线期DNA的合成,则联会复合体不能形成,联会不能进行,于是也不发生交换和交叉。
- 参考书目